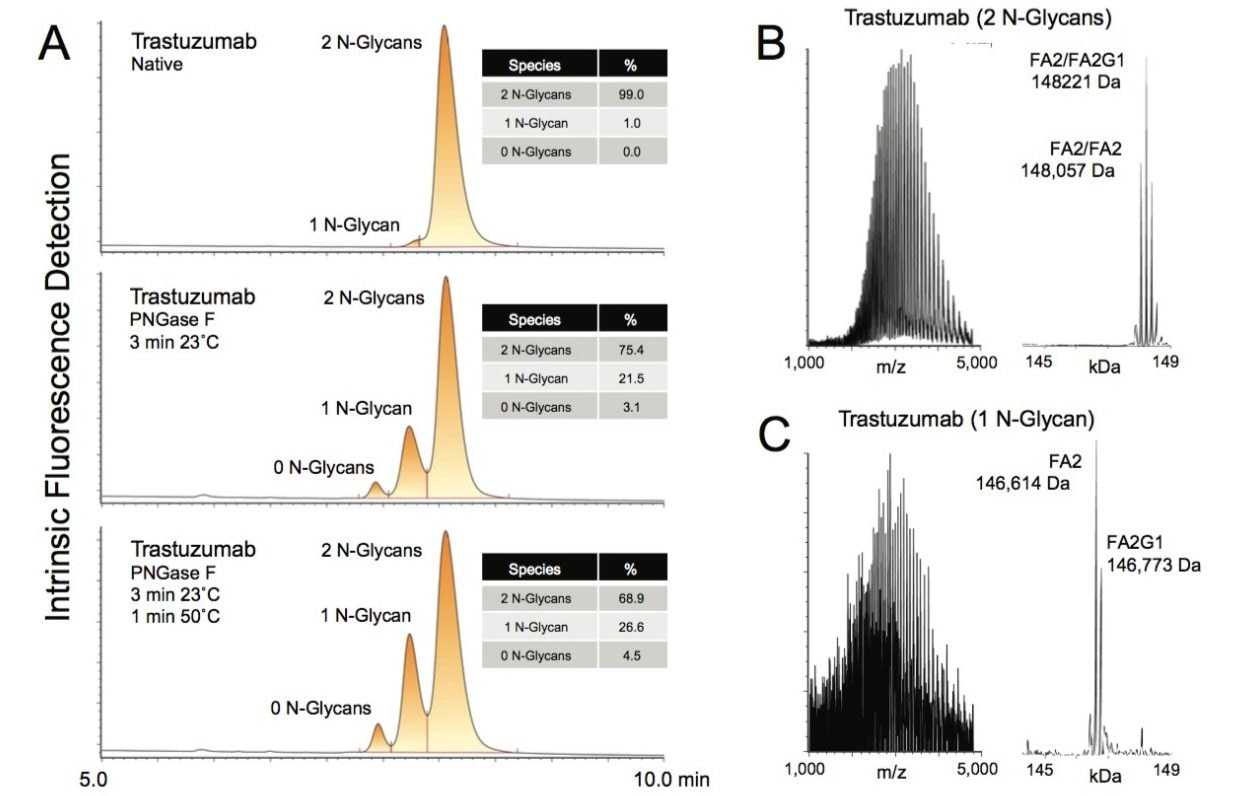
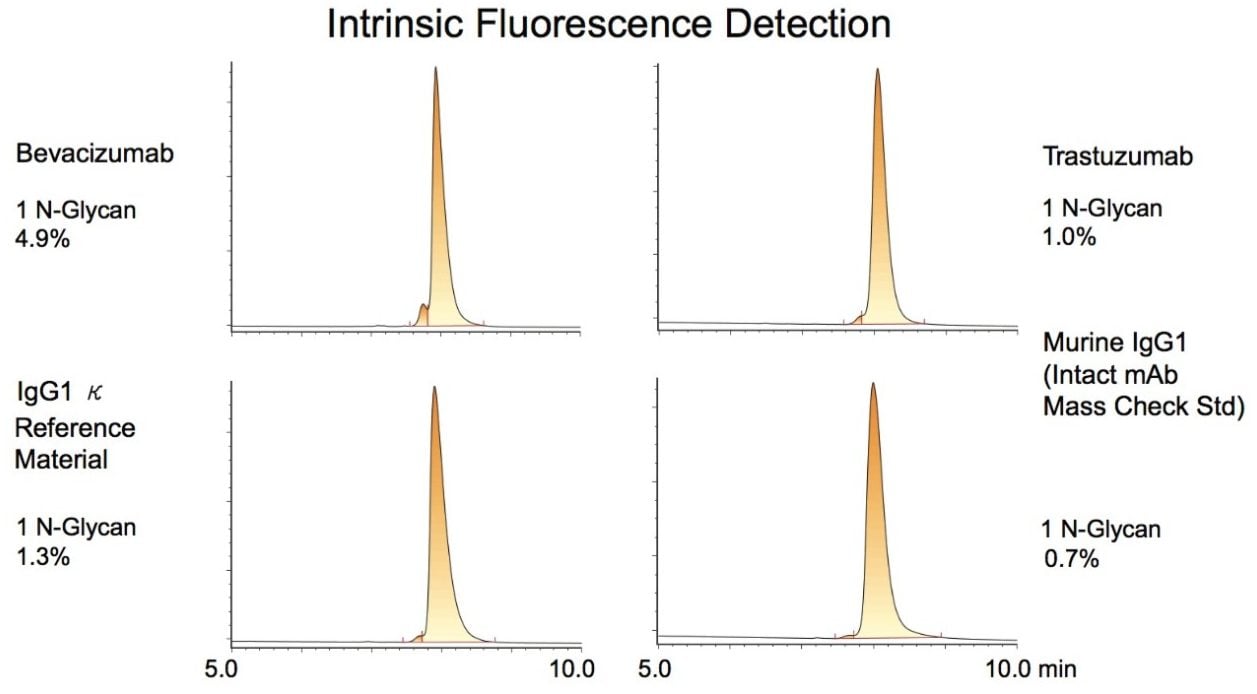
A high-throughput, high resolution HILIC separation was established for intact mAbs using a 2.1 x 150 mm wide-pore BEH amide column, a 0.4 mL/min flow rate, and a column temperature of 80 ˚C. In addition, two mobile phase additives, 0.1% trifluoroacetic acid (TFA) and 0.3% hexafluoroisopropanol (HFIP) were employed to improve the solubility of intact mAbs in the high organic, initial mobile phase conditions of the HILIC gradient. To enhance the sensitivity of this LC method, the intact proteins were detected by means of their intrinsic fluorescence. Excitation and emission wavelengths of 280 and 320 nm were found to provide optimal signal-to-noise and consistently flat chromatographic baselines that are desirable for peak integration. A representative set of chromatograms resulting from this 15 minute LC method is shown in Figure 1A. Three chromatograms are displayed. The top chromatogram shows trastuzumab as injected from a dilution of its formulation, while the other two chromatograms show samples of trastuzumab after being subjected to partial PNGase F deglycosylation. Deglycosylated samples of trastuzumab clearly showed three distinct peaks in their HILIC profiles, as was predicted if the glycan occupancy variants of a mAb were to be resolved. The unadulterated sample of trastuzumab contained measurable levels of only the fully occupied and singly occupied forms (1%) as confirmed by online mass analysis. The deconvoluted mass spectrum corresponding to the main LC peak exhibited several masses, such as 148,057 Da and 148,221 Da, that are within 2 Da of the theoretical molecular weights for the predominant glycoforms of trastuzumab (Figure 1B).9 In contrast, the deconvoluted mass spectrum for the smaller, less strongly retained peak showed reduced heterogeneity and masses that were lighter by approximately 1000 to 2000 Da, consistent with the single occupancy form (Figure 2B). Fully aglycosylated species were not detected in this sample. An interesting observation from these data is that the levels of fully deglycosylated forms are lower than would be predicted if both sites were deglycosylated at the same rate. In these examples, the observed level of fully deglycosylated forms is approximately one-third lower than would be predicted. This may indicate that either the digestion rate for one of the N-glycans is slower than the other, or that upon removal of the first N-glycan, the digestion rate of the remaining N-glycan is reduced.