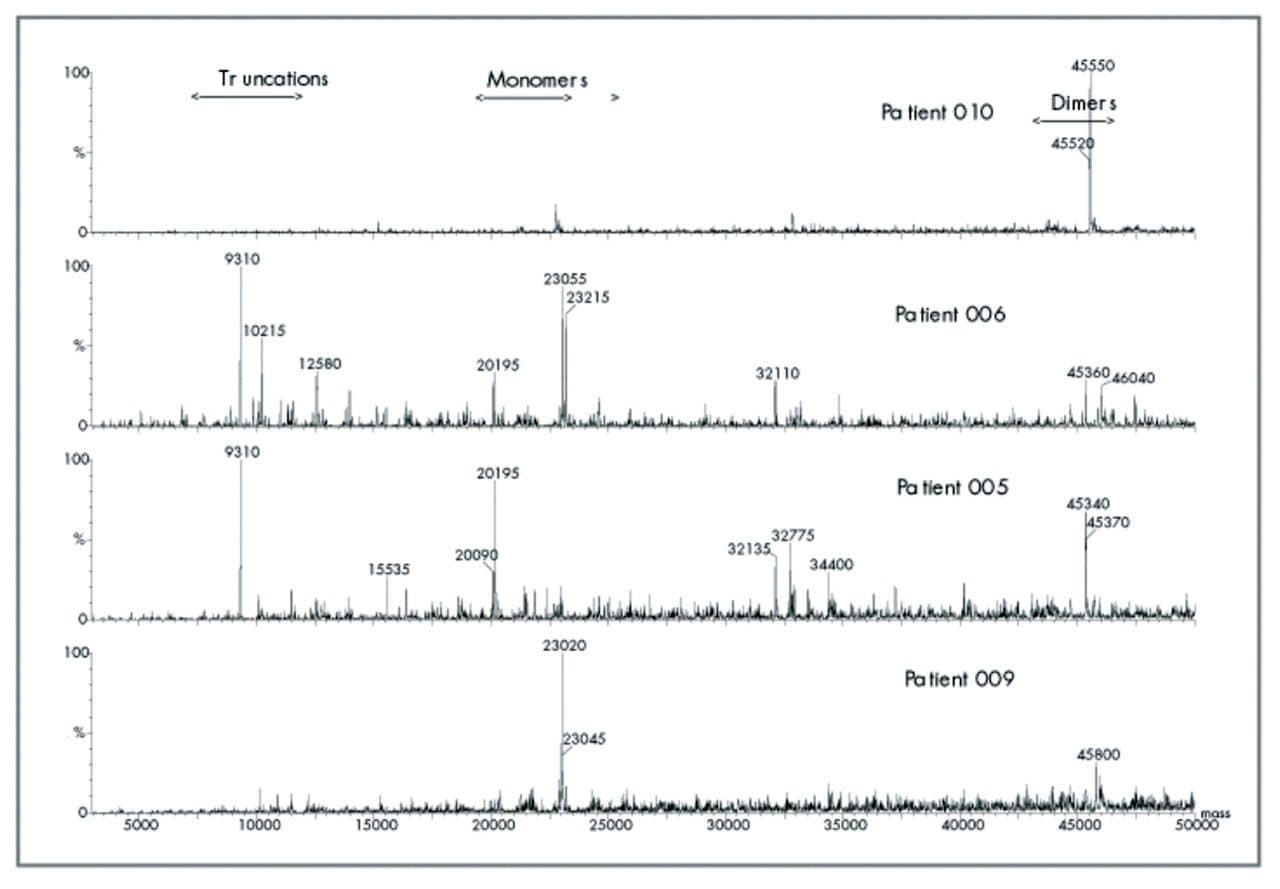
Procedure
Affinity column preparation
Polyclonal antibodies specific to free kappa and free lambda light chains (gift from Dr. A. R. Bradwell and The Binding Site, Ltd., Birmingham, England) were coupled to POROS AL media utilizing the "bulk" method as per the manufacturer's instructions. Approximately 2 mg of protein were coupled to 25 mg of affinity resin. Individual microbore guard columns (1 x 20 mm PEEK, Upchurch Scientific, Oak Harbor, WA) were packed with the anti-free kappa light chain or anti-free lambda light chain POROS media (~17 µL bed volume). Binding buffer was 10 mM phosphate pH 7.4, 150 mM NaCl. Elution buffer was 0.1M glycine pH 2.5. Bound light chain was washed with binding buffer for six minutes prior to elution with elution buffer (step gradient) for three minutes.
Protein trap/desalting column
A 1 x 20 mm PEEK microbore guard column packed with 40 µm CA silica (Bakerbond Prepscale WP Butyl (C4), J.T. Baker Research Products, Mallinckrodt Baker, Inc., 222 Red School Lane, Phillipsburg, NJ) was utilized to desalt the immuno-purified light chain prior to MS analysis. Elution of the desalted protein from this trap was via an organic gradient utilizing the C/D pumps. The gradient program was 2% D for three minutes (de-salts bound light chain), two minute linear gradient to 95% (elutes light chain), three minutes at 95% D, one minute linear gradient to 2% C (return to starting conditions).
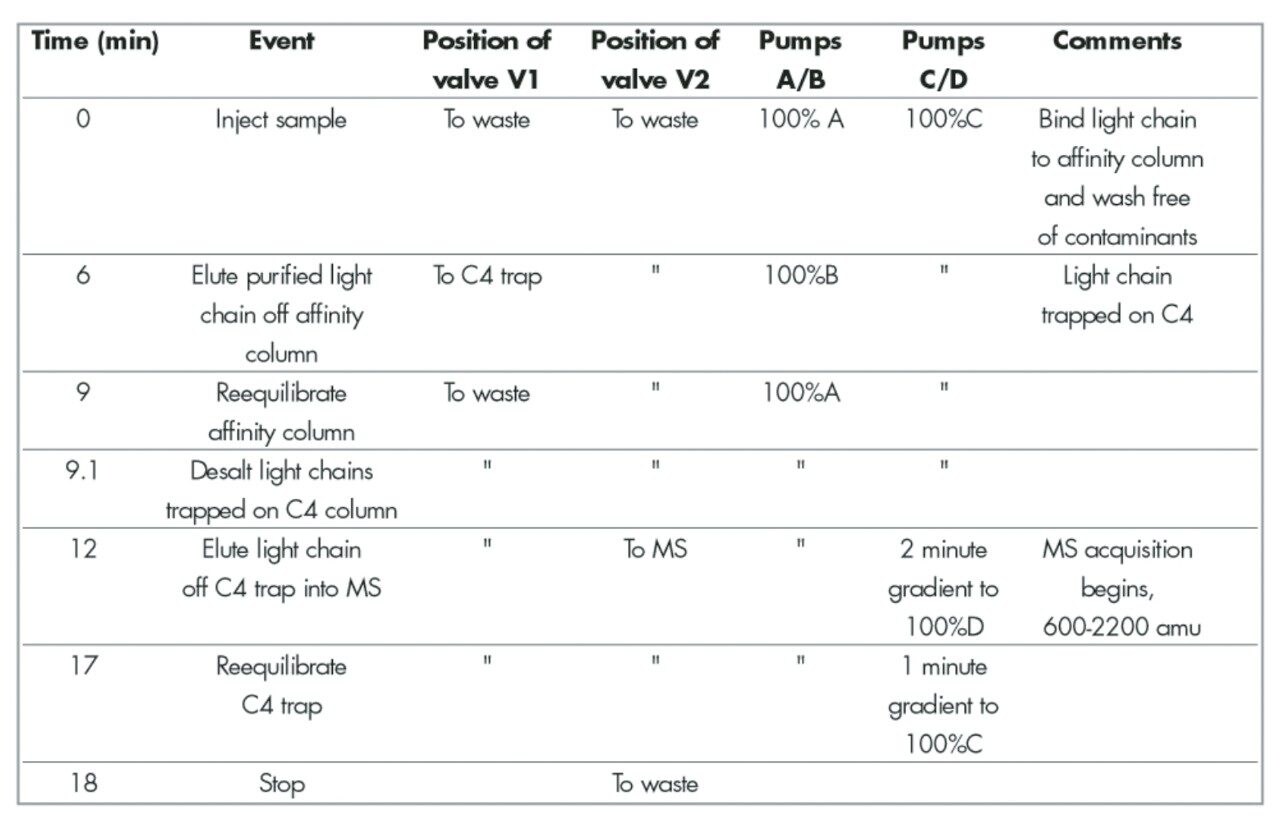
Event sequence
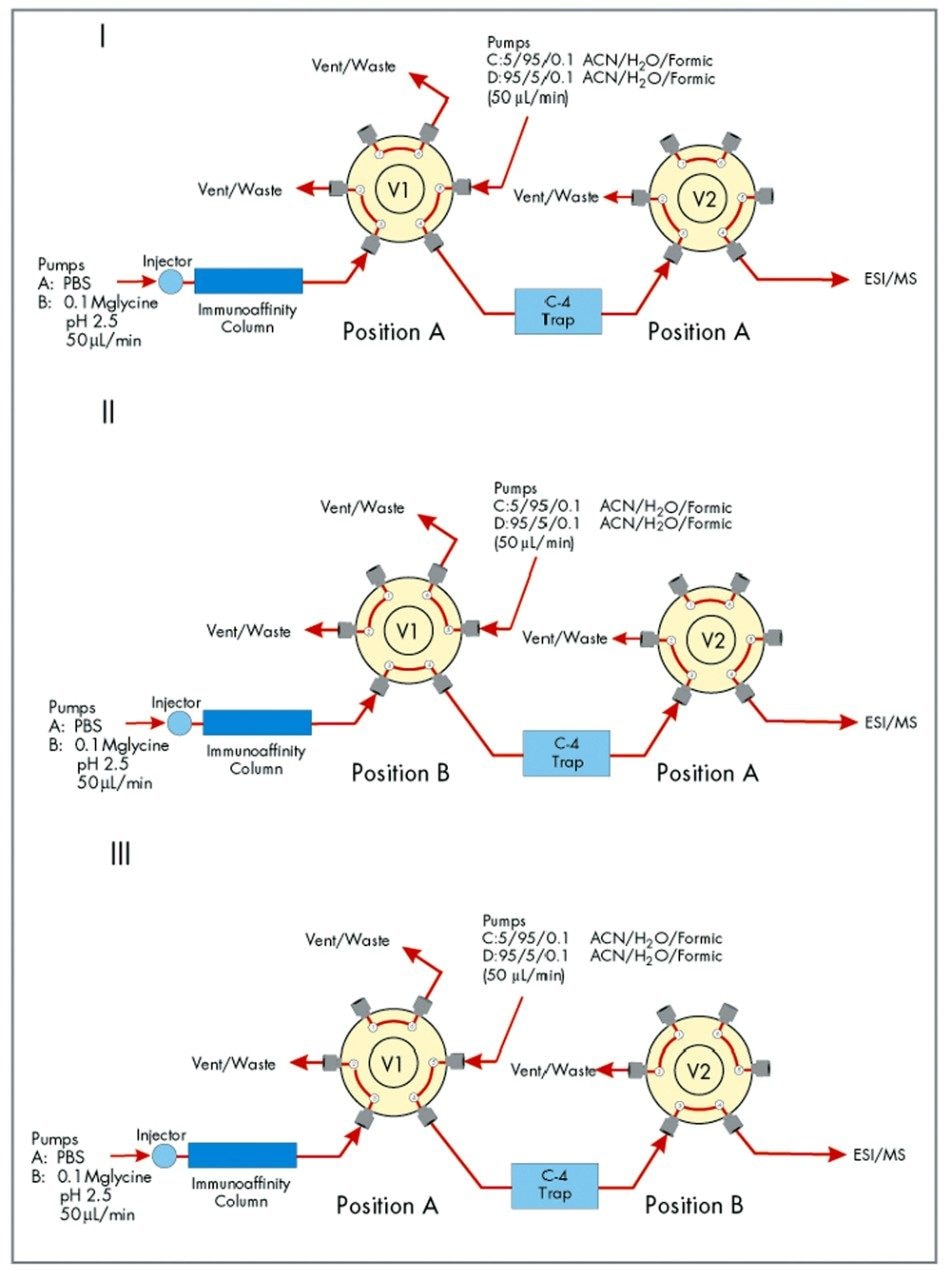
One two-position valve is positioned prior to the affinity column and is plumbed to switch between binding buffer (A: PBS) and elution buffer (B: 0.10 M glycine, pH 2.5). Another two-position valve V1 is plumbedto allow effluent from the affinity cartridge to flow to waste during the binding and washing periods and to the C4 trap during elution of bound light chain. The valve V2 is used as a divert valve to divert buffer salts from the ESI source.
Briefly, at time zero a sample is injected. Light chain binds to the affinity cartridge while valve V1 diverts unbound serum components to waste leaving light chain bound to the affinity cartridge. At t=6 minutes valve V1 diverts affinity column effluent onto the C4 trap. The light chain bound is released by switching to elution buffer B (6–9 minutes) and the eluted light chain is trapped on the C4 cartridge. Valve V1 returns (9 minutes) to its original position diverting the affinity cartridge effluent to waste allowing pumps C/D to pump across the C4 trap. Valve V2 diverts the first three minutes of the organic gradient (100% C, 9.1–12 minutes) to waste preventing any buffer salts from entering the source. At t=12 minutes valve V2 takes effluent from the C4 trap into the ESI source. A two minute gradient to 100% D (12–14 minutes) elutes light chain (~16 minutes) from the C4 trap. The gradient stays at 100% D (14–17 minutes) before coming back to 100% C (17–18 minutes).