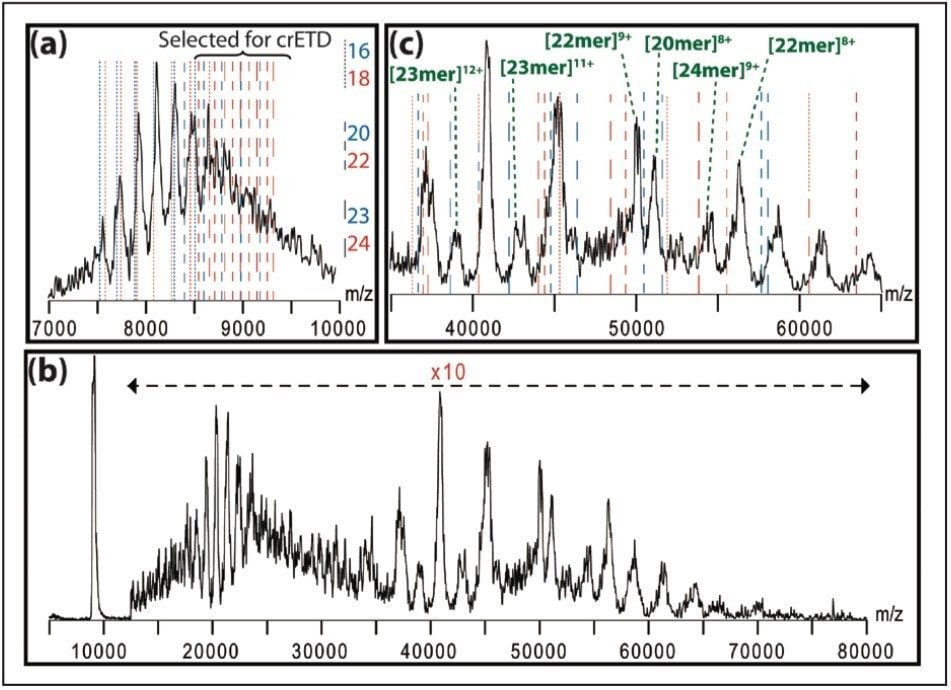
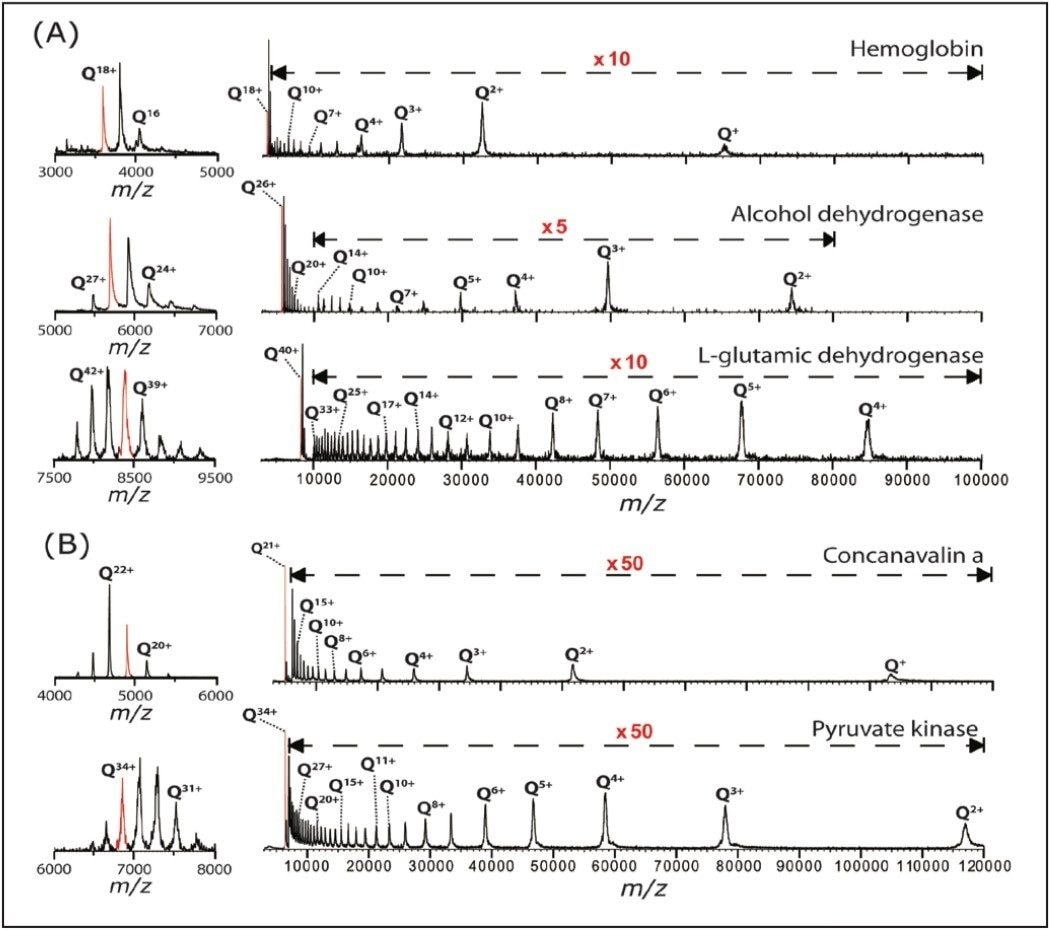
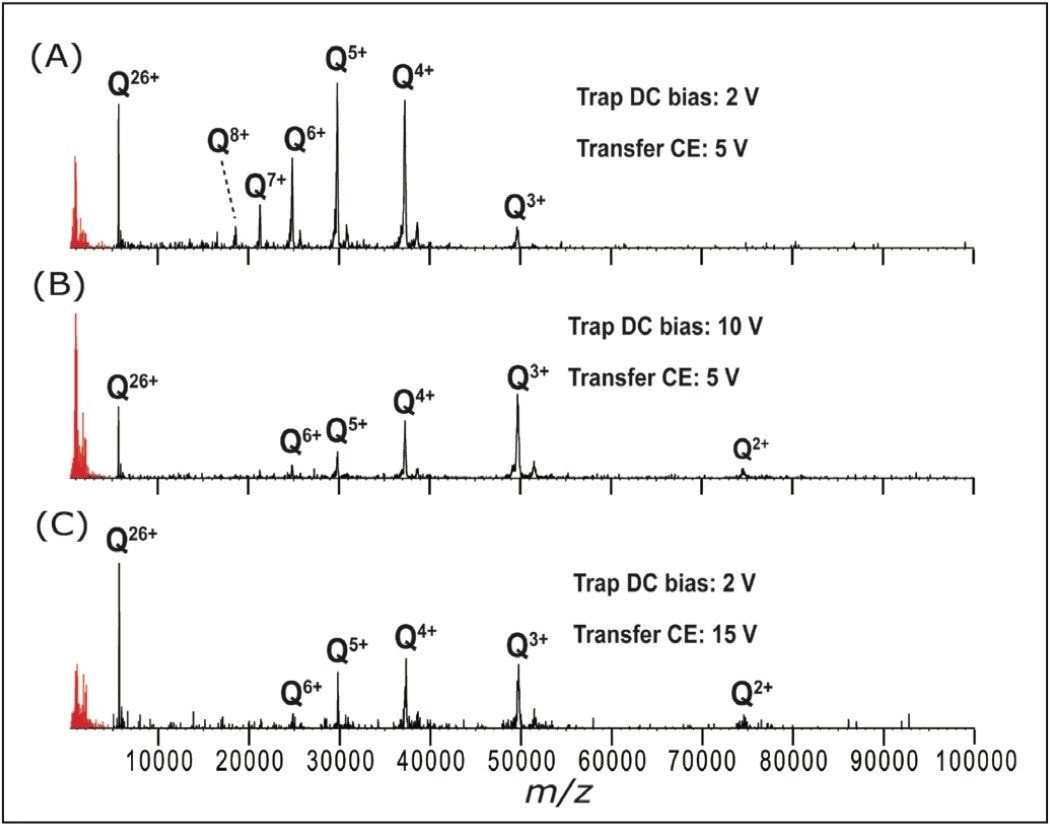
The production of high-mass, multiply charged ions via electrospray ionization (ESI) permits mass spectrometric analysis of native protein complexes.1,2 However, given the relatively broad peaks typical in native MS, mass and charge state assignment can be challenging, particularly for heterogeneous samples.3 Furthermore, a correlation exists between protein charge state and conformation, and lower charge states are often believed to resemble the native (solution) state more closely.4 Therefore, methods of reducing the gas-phase charge state offer analytical benefits. Previous efforts have generally either relied on ejection of highly charged monomers,3 causing significant disruption of the native structure, or have required instrument modification.5
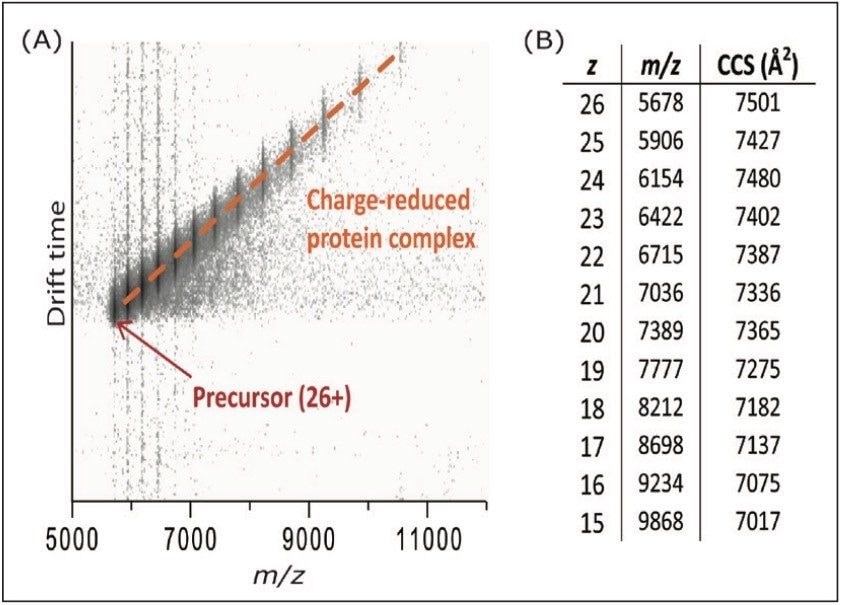
Here, we describe a method of extensive charge reduction of native protein complexes on unmodified SYNAPT G2 and SYNAPT G2-S Ion Mobility Mass Spectrometers. This results in increased peak separation and simplified spectral assignment, as illustrated in Figure 1, but also allows us to investigate electrostatic effects on protein folding state by using ion mobility. Concomitantly, some ETD fragments, originating from surface-exposed terminal regions, are also observed, providing sequence information.