The outbreak of the recent COVID-19 pandemic, caused by the SARS-CoV-2 coronavirus, has resulted in the sickening of millions and the deaths, to date, of over 2 million people globally.1 Because of its relative novelty, the scientific community has been engaged in rapid characterization and compilation of the biophysical characteristics of the virus, in hopes of aiding the development of effective vaccine and immunotherapies. The SARS-CoV-2 virion enters the cell by binding to the host cell ACE2 (angiotensin-converting enzyme 2) receptor with a protrusive transmembrane spike protein.2 The SARS-CoV-2 coronavirus spike protein is a homotrimeric class I fusion protein consisting of two heavily glycosylated subunits, S1 and S2.3,4 Previous studies of known viral pathogens, including influenza, have revealed the glycan composition of the outer envelope plays a key role in immunoevasion, notably through steric hindrance of immunorecognition sites.5–7
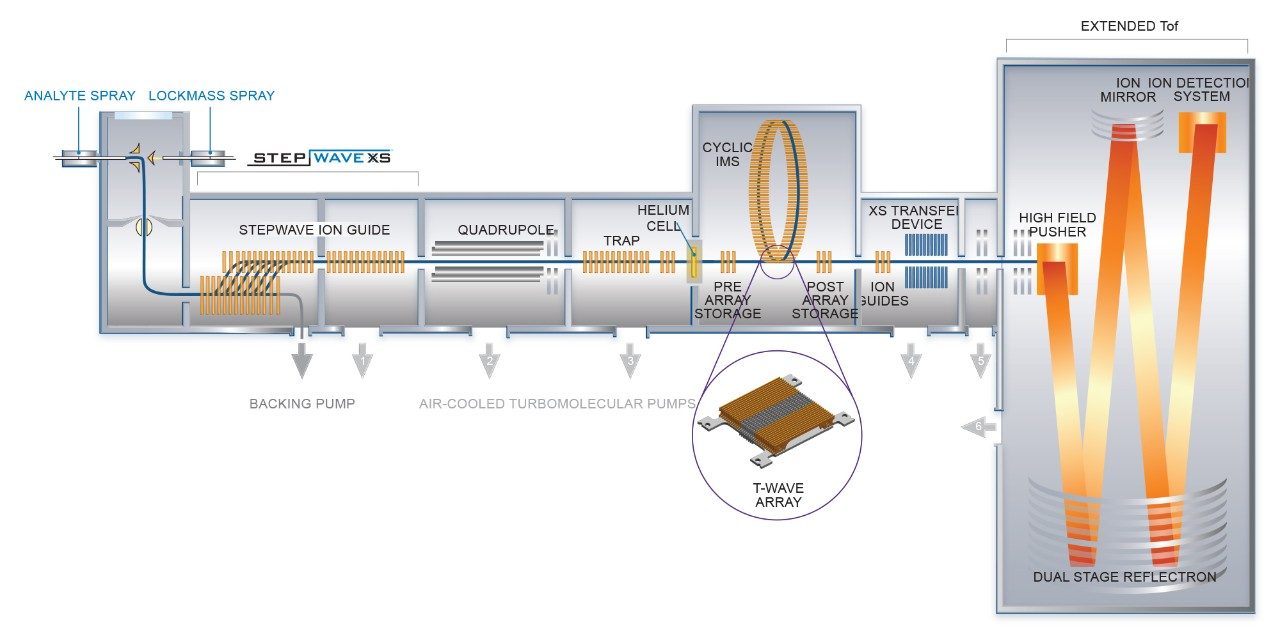
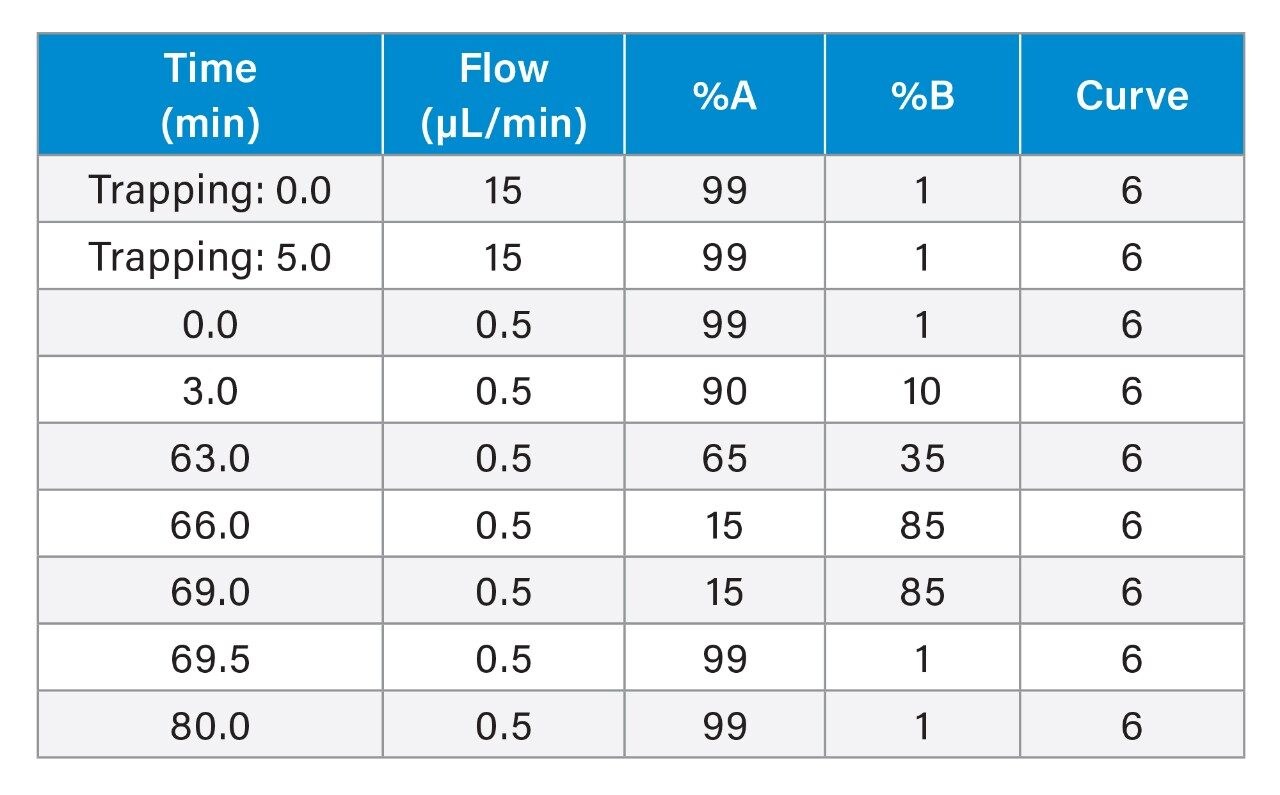
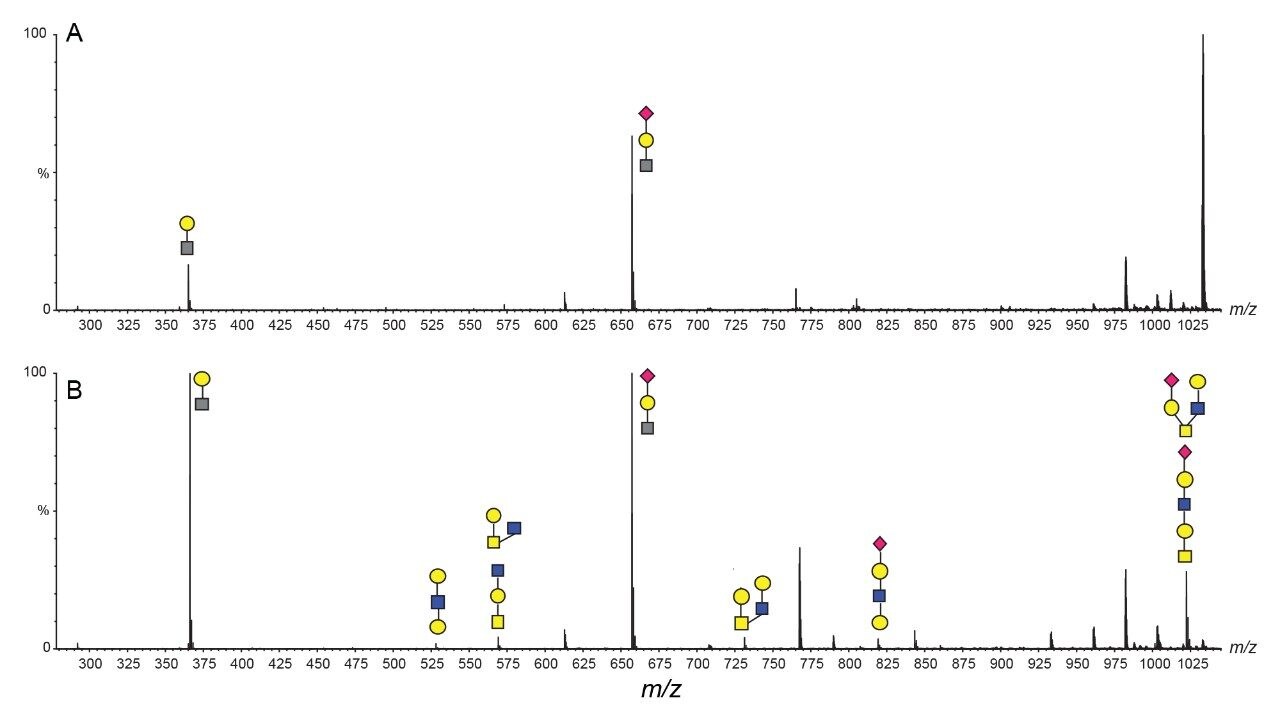
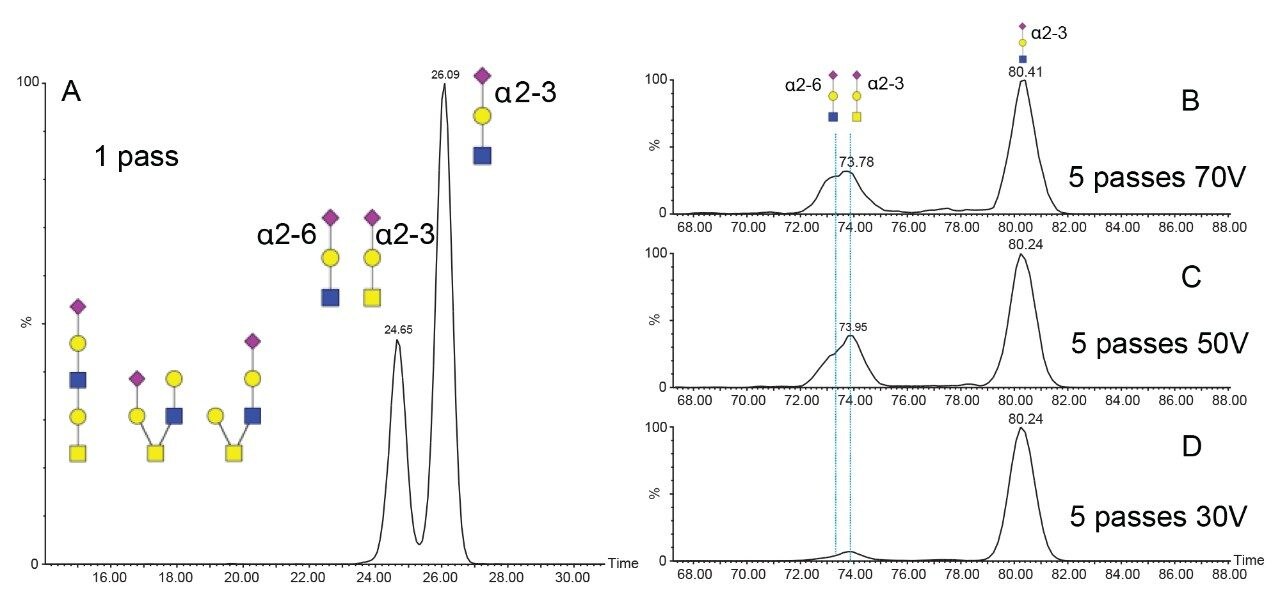
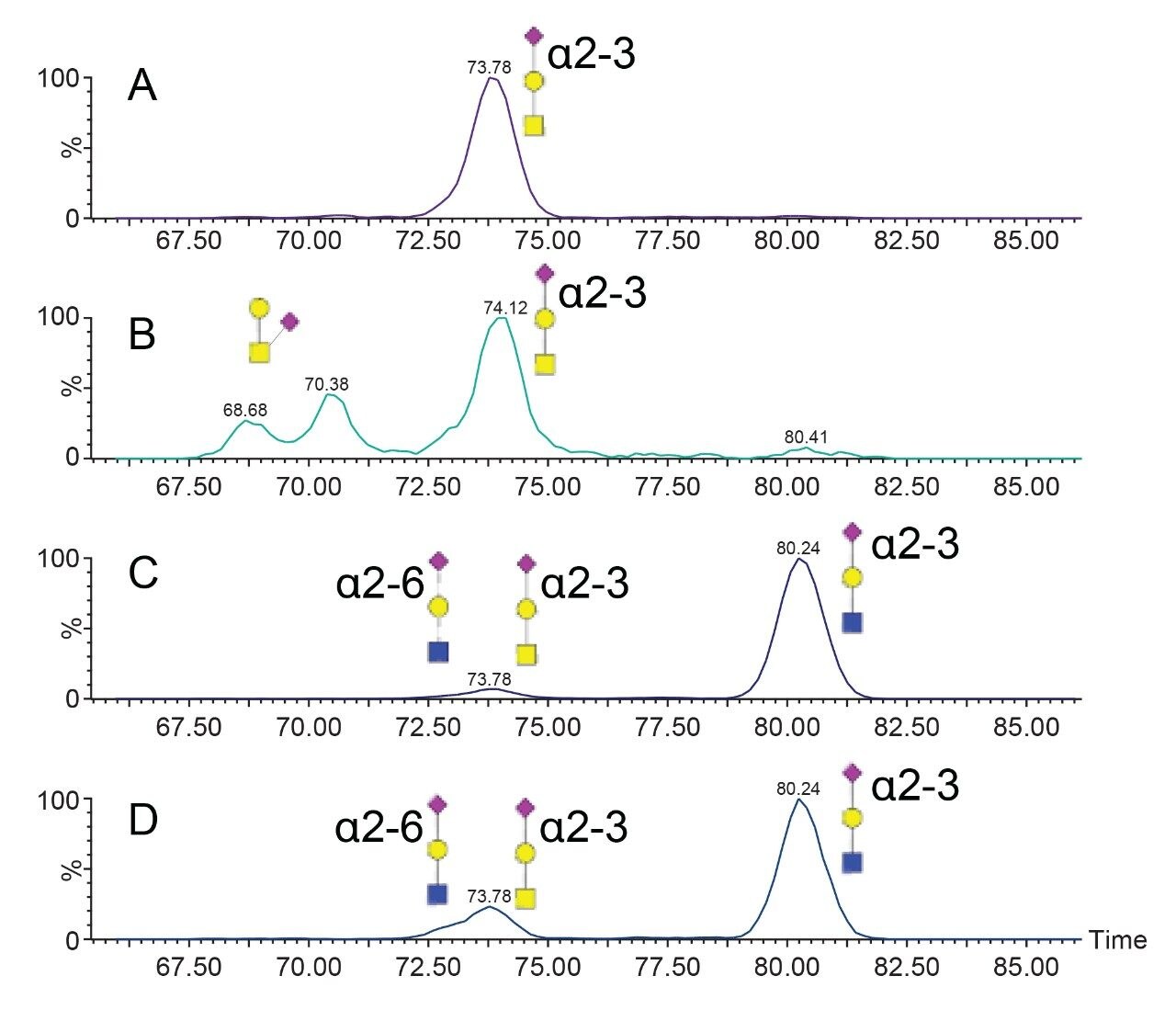
The characterization effort of the SARS-CoV-2 pathogen has included several comprehensive studies of the glycosylation of the coronavirus spike protein. To date, these studies have indicated consensus occupancy of 14 of the 22 available N-linked glycosylation sites and conflicting occupancy of the remaining seven sites.8–16 In addition, Shajahan et. al showed evidence for three O-linked glycosylation sites on the S1 domain, one of which was confirmed by Sanda et. al.10,11 In a sequence analysis of the SARS-CoV-spike protein, Andersen and co-workers predicted a furin cleavage site, a site critical to protein activation, in the linker region flanking the S1 and S2 subunits.2 This region containing the furin cleavage site was further predicted to contain up to three additional O-linked glycosylation sites, the function of which was speculated to be related to infectivity and transmissibility.2 Characterization of O-linked glycans is notoriously difficult for several reasons: the lack of a consensus sequence, high heterogeneity among the glycoforms, and low relative abundance. Consequently, characterization of these types of analytes requires high performance chromatography and high resolution, high sensitivity mass spectrometry. Previous studies using the Waters SYNAPT platform have shown the capacity to resolve branched and linear glycan structures of isomeric glycans and separate α2-3 and α2-6 linkage isomers from glycan standards.12–16 MS/MS fragmentation with the Waters SELECT SERIES Cyclic IMS provides clear evidence of O-linked glycosylation along the linker region preceding the polybasic furin cleavage site and cyclic ion mobility is used to site-specifically separate and characterize the O-linked glycan structures. The scalable resolution afforded by this unique experiment provides evidence for a mixture of core 1, extended core 1, and core 2 structures, with resolution of NeuAcα2-3Galβ1-3GalNAc and NeuAcα2-6Galβ1-4GlcNAc, isomers that feature highly similar collisional cross sections.