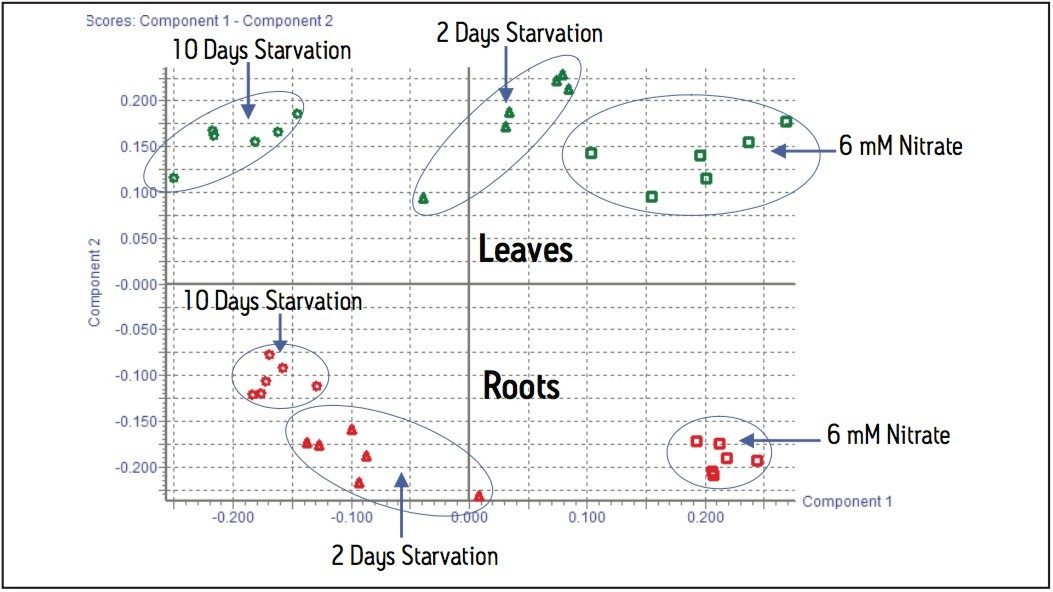
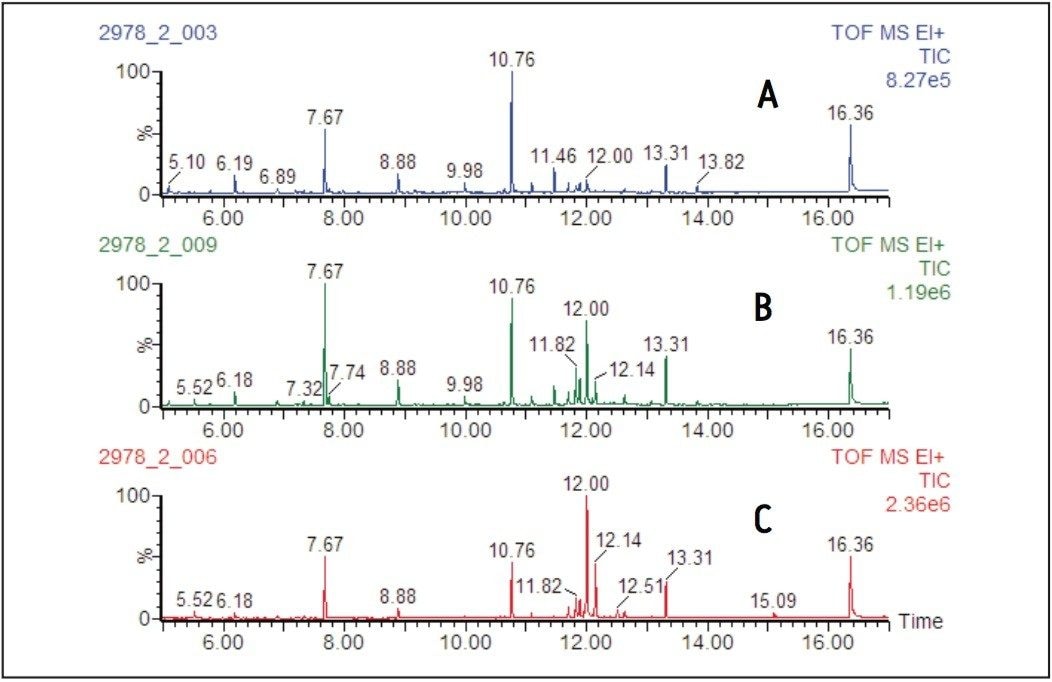
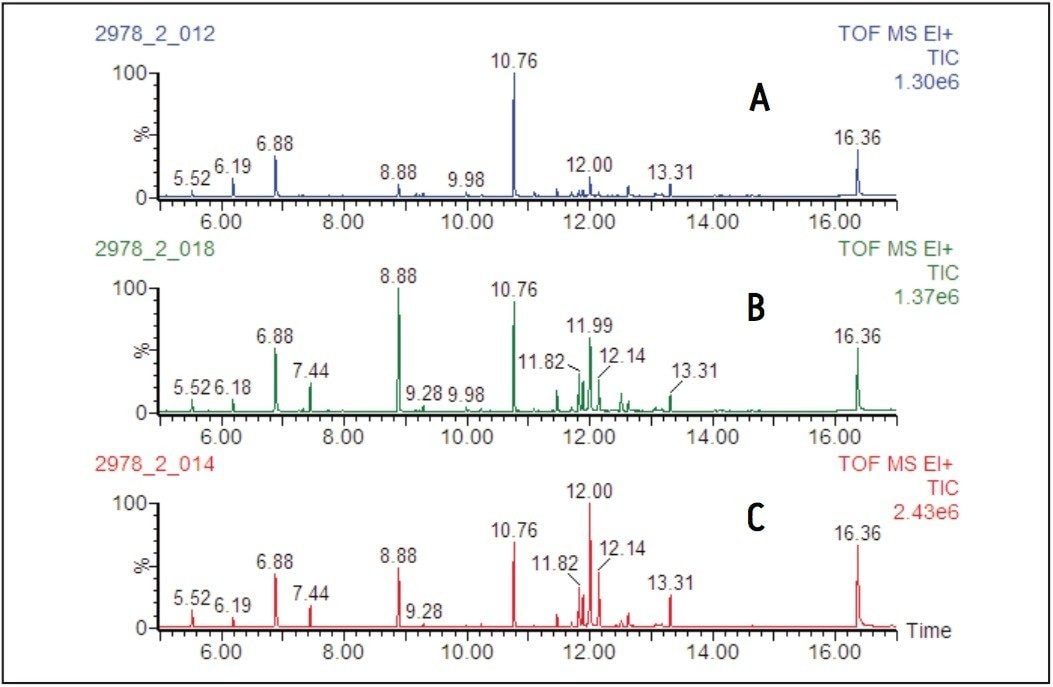
Examination of the chromatograms shows distinct differences between the metabolic profiles of the leaves and roots, with the major peak at 7.67 minutes in the leaf samples, assigned as fumaric acid, only being present at trace levels in the root samples.
Conversely, in the root samples the peak at 6.88 minutes, mainly composed of tris(trimethylsilyl) phosphate, is only present at a low level in the leaf samples.
The major changes observed as a result of the nitrogen starvation, however, were an increase in intensity of the peaks in the retention time range of 11.75 to 12.25 minutes, corresponding to an increase in carbohydrates.
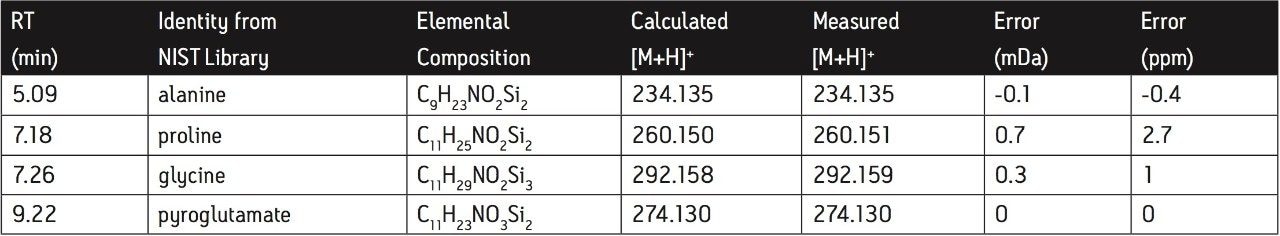
The levels of amino acids were observed to decrease during the time course of the nitrogen starvation, consistent with the results reported by Urbanczyk-Wochniak and Fernie.2 The amino acids were identified using the NIST library of EI spectra and their molecular masses confirmed by exact mass CI analysis.
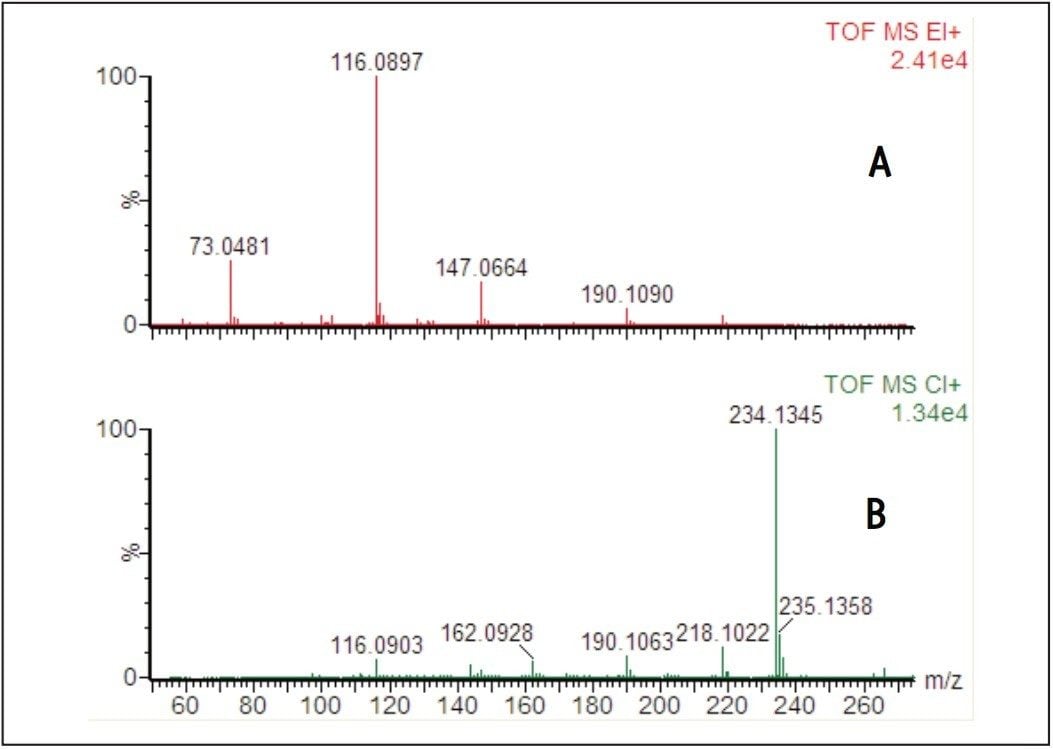
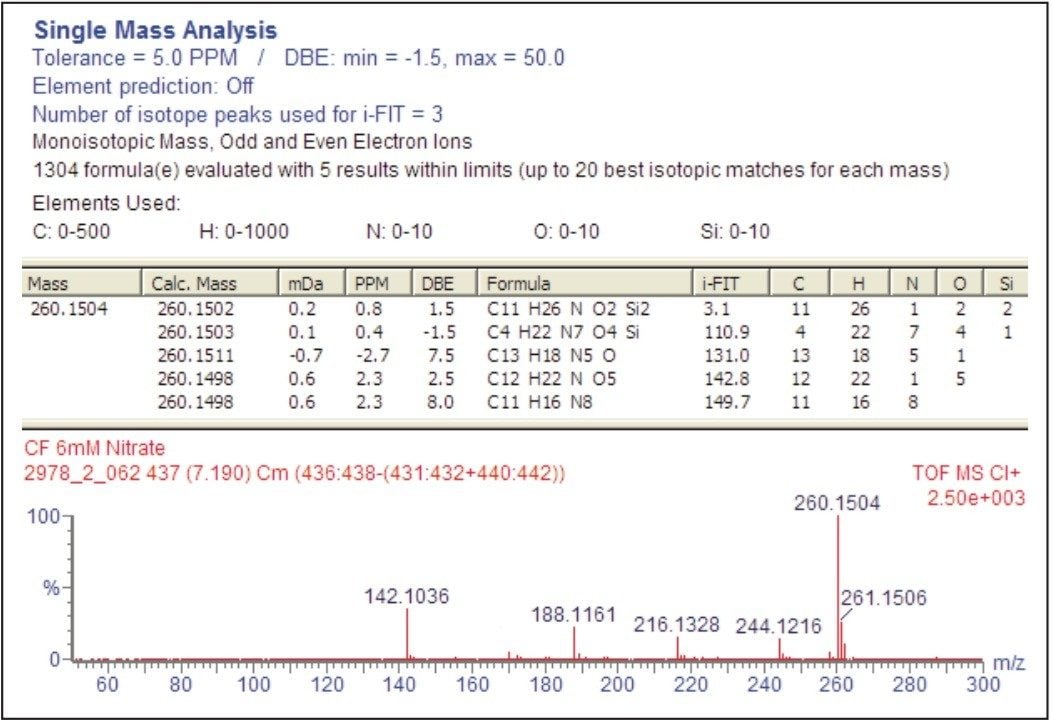
Representative EI and CI spectra obtained from a leaf sample grown under conditions of adequate nitrogen for plant growth are shown in Figure 5. The EI spectrum gave a good match for the bis-trimethylsilyl derivative of alanine and the molecular mass was confirmed by exact mass CI with a measured mass of 234.1345 (error -0.1 mDa, -0.4 ppm) for the protonated molecule.