Post-translational modifications (PTMs) and chemical degradations impact the efficacy and safety of therapeutic proteins and peptides. Therefore, they have to be effectively controlled and monitored during production, formulation, and storage. Liquid chromatography (LC)-based peptide mapping with ultra violet (UV) and/or mass spectrometry (MS) detection is a method of choice for characterization of PTMs and degradations.1
However, higher throughput and more sensitive methods are needed for routine monitoring and quantification of protein drugs during development and manufacturing. For example, in protein formulation development hundreds of samples are typically analyzed to insure purity and stability. Although intact protein mass measurement (including limited proteolysis followed by LC-MS analysis) can be used for fast evaluation of large mass-shifted PTMs such as glycosylation, it does not provide information on modification sites and has limitations for detection of modifications with small mass shifts,2, 3 such as asparagine (N)-deamidation, aspartic acid (D)-isomerization, and methionine (M)-oxidation.
Monoclonal antibodies (mAbs) are an important class of biotherapeutics. They are susceptible to a number of PTMs and degradations common to other classes of therapeutic proteins. Peptide mapping studies3, 4 demonstrate that PTMs and degradations, such as glycosylation, deamidation, and oxidation, only occur in specific motifs of a mAb. This application note evaluates the use of targeted UPLC-MS analysis using multiple reaction monitoring (MRM) for fast and sensitive monitoring and quantification of degradations in mAb digests.
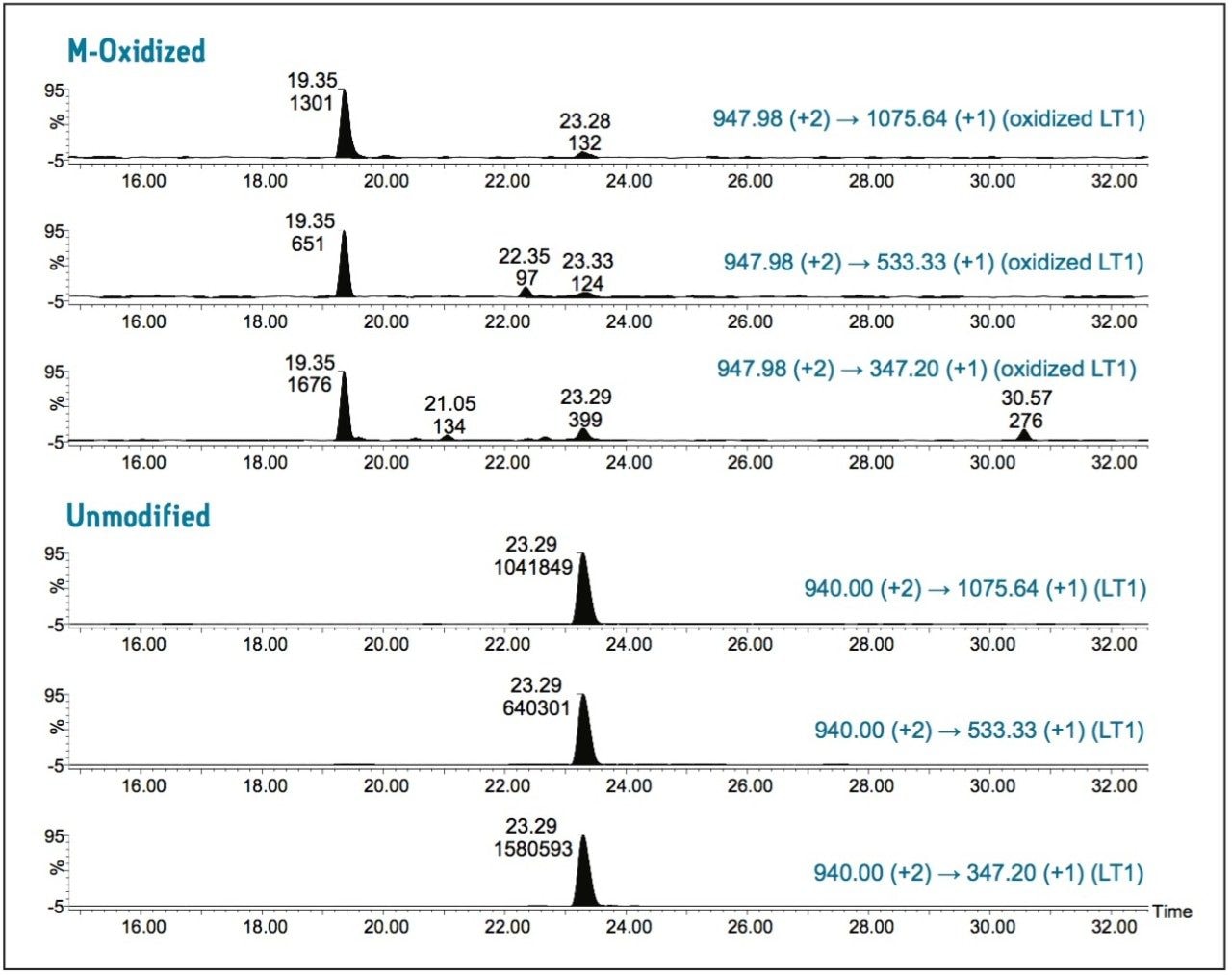
As a proof of concept, we performed a UPLC/MRM study with the goal of confirmation and quantification of site-specific M-oxidations in a recombinant IgG1 tryptic digest previously identified by UPLC/MSE peptide mapping studies.3, 4
Both oxidized and unmodified M-containing tryptic peptides were unambiguously determined by multiple MRM transitions, and relative quantification data were obtained for a low-abundance M-oxidized peptide.
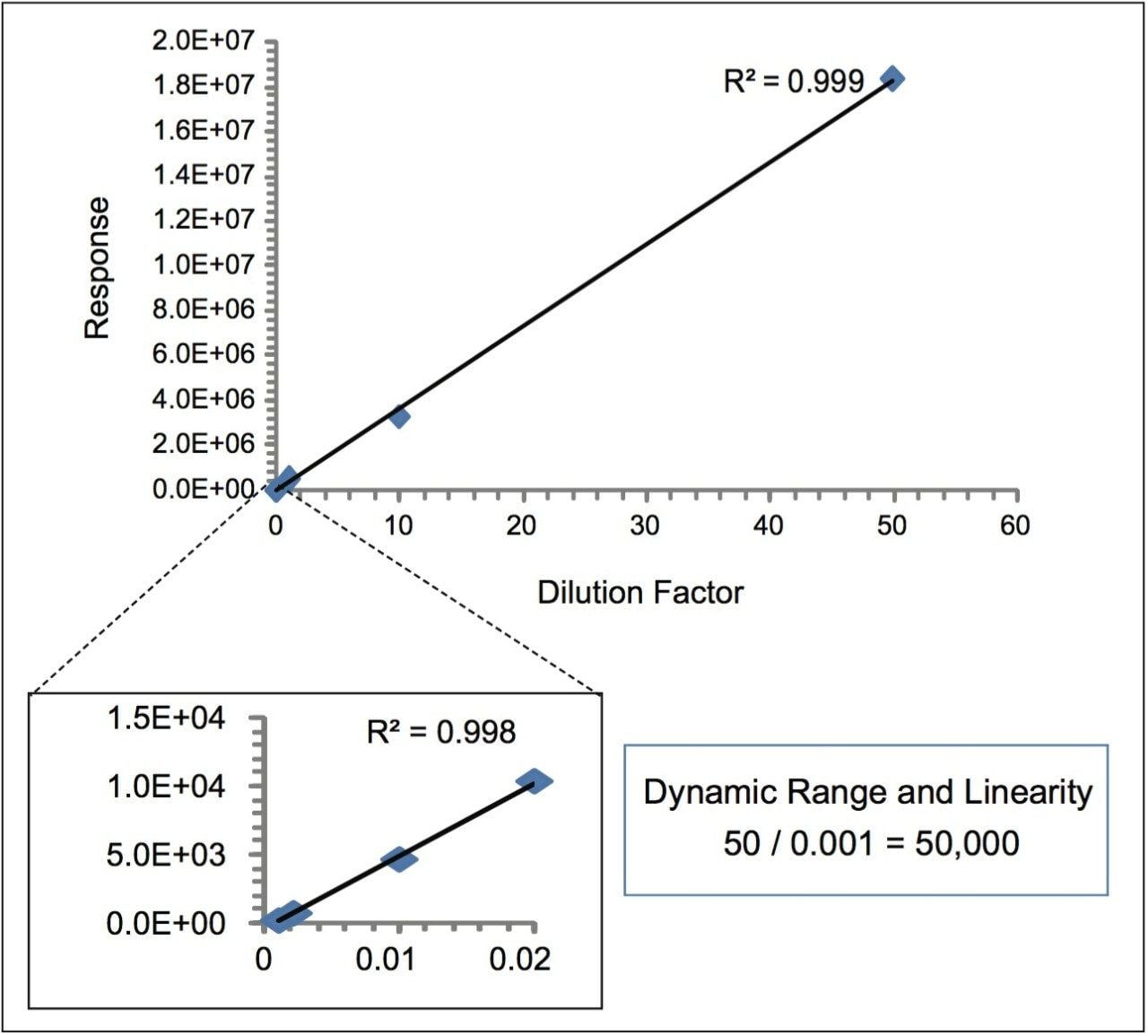
The large dynamic range and linearity of the combined ACCQUITY UPLC/Xevo TQ MS System5,6 used for this UPLC/MRM assay offers a significant improvement in speed, specificity, and sensitivity of mAb analysis and has the potential to expedite the research and development of protein drugs by saving both time and analytical resources.