Multiple Quality Attribute Characterization of a Ribonucleoprotein Complex by SEC-MALS
Katayoun Morakabia, Sophia Kenricka, Matthew Gardnerb
aWaters Corporation, United States
bScribe Therapeutics, United States
Published on June 12, 2026
Abstract
Size-exclusion chromatography coupled with multi-angle light scattering (SEC-MALS) is a powerful solution for analysis of ribonucleoprotein (RNP) complexes and their individual components, essential for the development of potent and consistent molecules. The study presented here highlights a SEC-MALS method that leverages the ASTRA™ Software ADC Analysis Module to quantify ribonucleoprotein (RNP) composition (molar mass and fraction of clustered regularly interspaced short palindromic repeats (CRISPR)-associated protein (Cas) protein and single guide RNA (sgRNA)) at each point across the chromatogram and determine RNP stoichiometry under native conditions. The SEC-MALS data revealed the presence of multiple RNP complexes with the most abundant complexes containing a non-stoichiometric number of sgRNA molecules. The data suggests that the oligomeric state of the RNP complex is dependent on the oligomeric state of the sgRNA. The approach presented here provides robust characterization of CRISPR/Cas components and supports researchers in improving RNP complexation.
Benefits
- Detection of aggregation and higher-order species in RNP components including free sgRNA and CasX
- Quantification of RNP composition and stoichiometry under native conditions
- High-performance separation enabled by the XBridge™ Premier Protein SEC 250 Å Column
Introduction
CRISPR and CRISPR-associated protein (Cas) systems were originally discovered as an adaptive immune mechanism, conferring resistance against bacteriophage through sequence-specific cleavage of viral DNA. Over the last two decades, CRISPR/Cas systems have been applied as an innovative gene editing tool in mammalian cells.1 The CRISPR/Cas system comprises a Cas nuclease and a guide RNA (gRNA) which form a rRNP complex that targets and cleaves DNA at specific loci. The gRNA directs the nuclease to the target sequence where the enzyme subsequently binds and cleaves DNA, creating double-stranded breaks.
Double-stranded breaks (DSBs) generated by Cas are repaired by endogenous cellular pathways. One such pathway is the homology-directed repair (HDR) pathway, where a homologous DNA sequence is used to repair the damaged DNA. For gene editing, scientists leverage this process by supplying an exogenous repair template that maintains homology to the target gene but carries desired sequence modifications, enabling precise genomic edits.2 Notably, CRISPR technology was applied in the first FDA-approved therapy for treatment of sickle cell disease and holds significant promise for addressing the underlying cause of genetic disorders by correcting the specific DNA sequences responsible for morbidity and mortality.3,4
Effective genome editing starts with the design and quality CRISPR/Cas components (Cas and sgRNA). Screening of the starting materials is crucial to the formation of an active RNP complex. Quality attributes (QAs) for the apoenzyme and sgRNA include native oligomeric state and purity (process contaminants and aggregates). LC-MS is widely adopted to determine identity and to monitor sgRNA sequence modifications. However, it may not be able to quantify aggregated species in solution. SEC under native conditions is commonly used to determine oligomeric state and quantify aggregates, fragments, and other species in solution. However, due to vast differences in conformation between the enzyme and sgRNA, the elution time often does not correlate with molar mass. SEC-MALS overcomes these challenges by quantifying the absolute molar mass of the desired product, aggregates, fragments, and other impurities under native conditions.
After ensuring the quality of the individual CRISPR components, quantifying and monitoring the composition of RNP complex is crucial to producing potent and consistent molecules. Although techniques such as native PAGE and gel shift assays can confirm that the sgRNA binds to the Cas protein, they are primarily qualitative and do not provide enough information about complex stoichiometry. Native mass spectrometry is a powerful technique widely used to characterize proteins and nucleic acids, but volatile buffers necessary for native MS can mask aggregates. LC-UV can be employed under native conditions and is used to assess RNP formation and purity; however, it does not directly measure molar mass or quantify the composition, which can lead to incorrect identification of the species in the solution.
This application note illustrates how SEC-MALS with UV detection at 260 nm and 280 nm and differential refractive index (dRI) detection can be used to assess multiple quality attributes of a CRISPR/Cas system in a single assay. This method enables quantification under native conditions, preserving the intact RNP complex as well as any potential aggregates for more robust characterization. First the native oligomeric state and purity of the free sgRNA and apoenzyme were evaluated. Then, SEC-MALS was applied to the RNP complex to directly measure the absolute stoichiometry of all the complexes present in solution, giving deeper insight into the heterogeneity of these molecules, which can ultimately impact their functionality. This app note highlights analytical tools for characterizing a CRISPR/Cas system and supports researchers to develop strategies to improve RNP complexation.
Experimental
Sample Preparation
sgRNA, CasX apoenzyme, and pre-assembled RNP complex were prepared by Scribe Therapeutics. The CasX apoenzyme has an expected molar mass of 110 kDa, and the corresponding sgRNA is expected to be 35 kDa.
LC Conditions
|
LC system: |
Waters Arc™ Premier HPLC System with a Quaternary Solvent Manager (QSM) |
|
Detection: |
Waters Arc Premier 2998 UV Detector, DAWN™ MALS Photometer, and an Optilab™ Differential Refractive Index (dRI) Detector |
|
Vials: |
LCGC Certified Clear Glass 12 x 32 mm Screw Neck Vial, Total Recovery, with Cap and Preslit PTFE/Silicone Septum, 1 mL Volume (p/n: 186000385C) |
|
Column(s): |
Waters XBridge Premier Protein SEC Column, 250 Å, 2.5 µm, 7.8 x 300 mm (p/n: 186009962) |
|
Column temperature: |
25 °C |
|
Sample temperature: |
4 °C |
|
Injection volume: |
All samples were injected neat, with injection amounts ranging from 15 µL to 45 µL |
|
Flow rate: |
0.5 mL/min |
|
Seal wash: |
20% HPLC-grade isopropanol / 80% Milli-Q® water (v/v) |
|
Mobile phase: |
50 mM sodium phosphate, pH 7.2, and 300 mM sodium chloride |
Data Management
|
Chromatography software: |
HPLC CONNECT™ 4 |
|
Data analysis software: |
ASTRA™ 8 Software with ADC Analysis Module |
Results and Discussion
Analysis of Free CasX and sgRNA
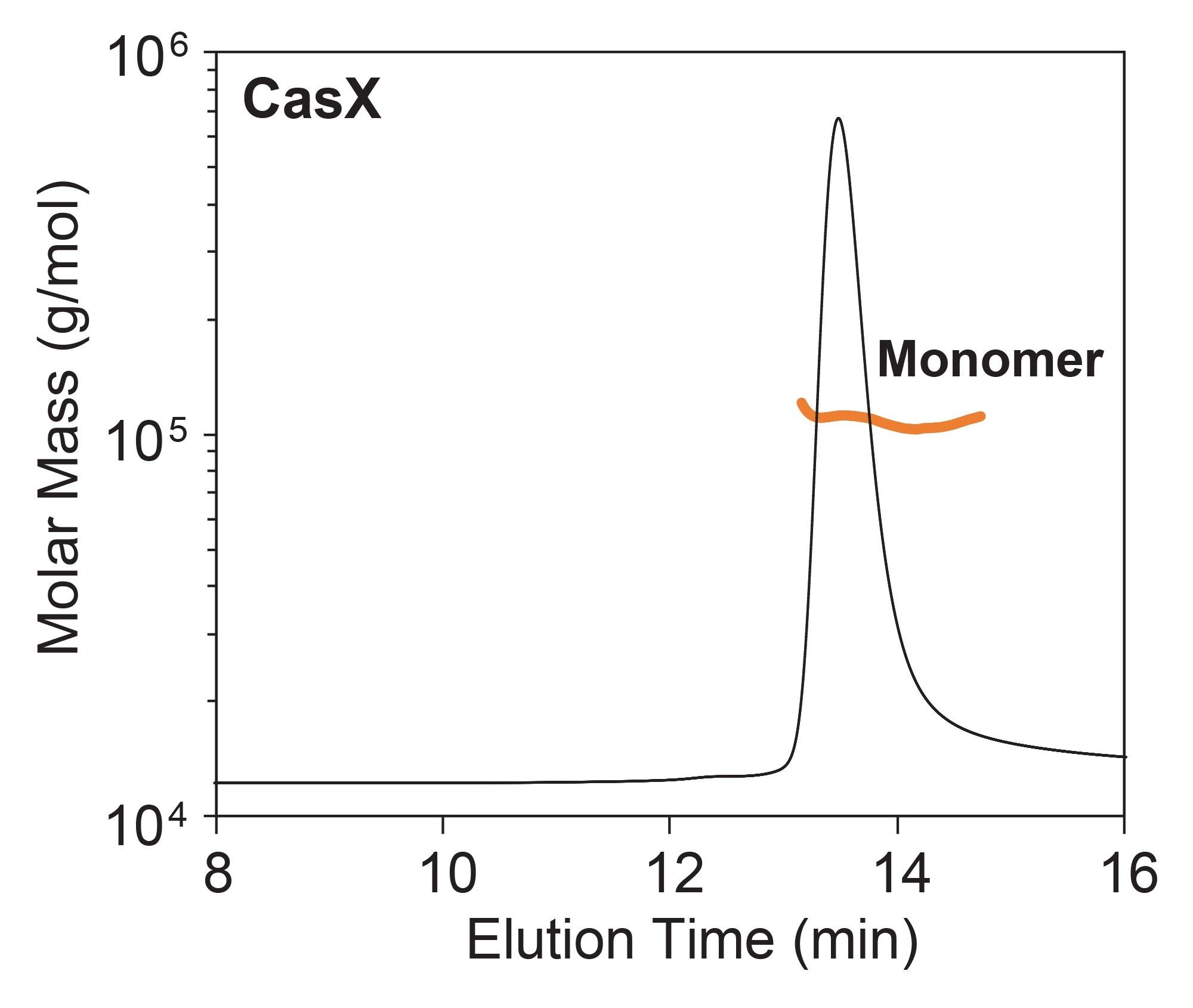
SEC-MALS of the CasX apoenzyme revealed that it is predominantly monomeric with a molar mass of 108 ±3 kDa (Figure 2), consistent with the expected molar mass. The molar mass distribution reveals a largely monodispersed sample which is favorable for RNP complexation. A minimal amount of (<2% by mass) of dimer was also detected.
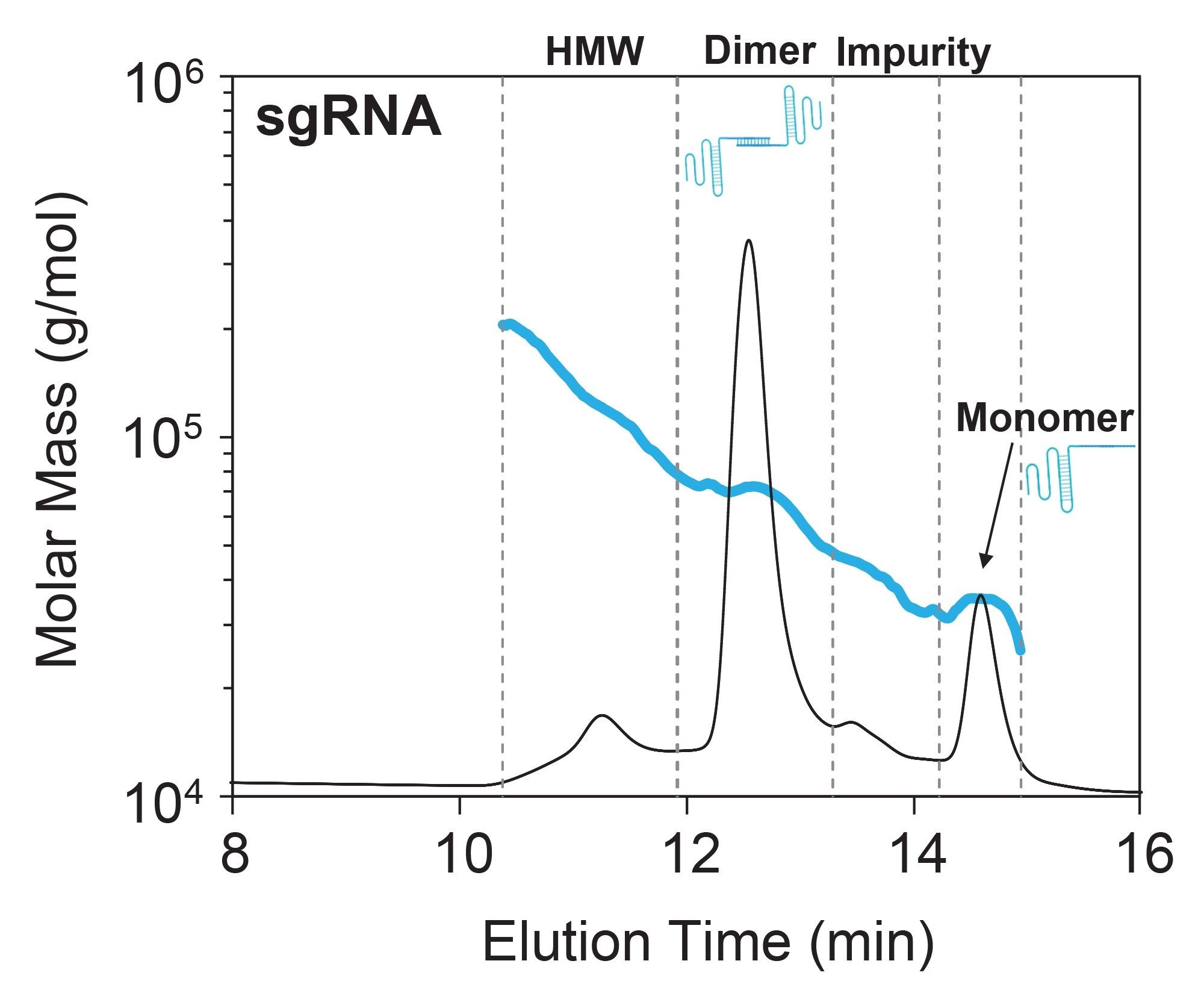
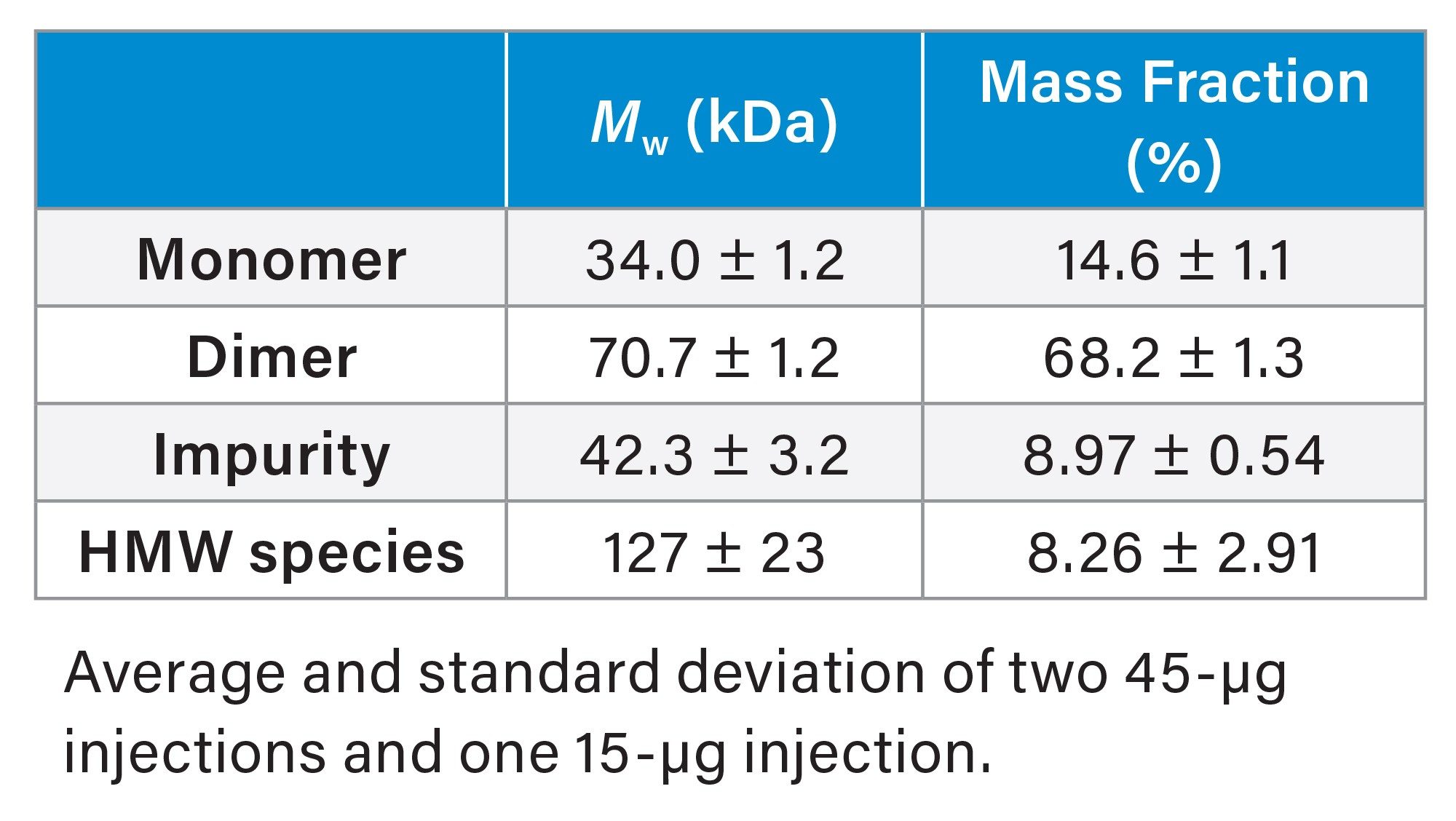
Unlike the mostly monomeric CasX protein, SEC-MALS of free sgRNA revealed significant higher-order structures (HOS) and aggregation, which could directly influence RNP complexation, heterogeneity, and DNA cleavage.5 Although the sgRNA monomer was clearly present, the dimer species made up ~70% of the eluted mass (Figure 3 and Table 1). This is not surprising given that sgRNA is designed with many self-complementary nucleotides which can fold into various secondary structures and HOS. Although the exact structure of this dimeric species is unknown, it is hypothesized to arise from Watson-Crick base pairing between two single-stranded sgRNA, forming partial double-stranded structures.6
In addition to the dimer, two other species were identified: a high molecular-weight (HMW) species and an impurity (<10% by mass). Both peaks exhibit a heterogeneous molar mass distribution suggesting mixtures of coeluting species (Figure 3).
Molar Mass and Stoichiometry of RNP Complex
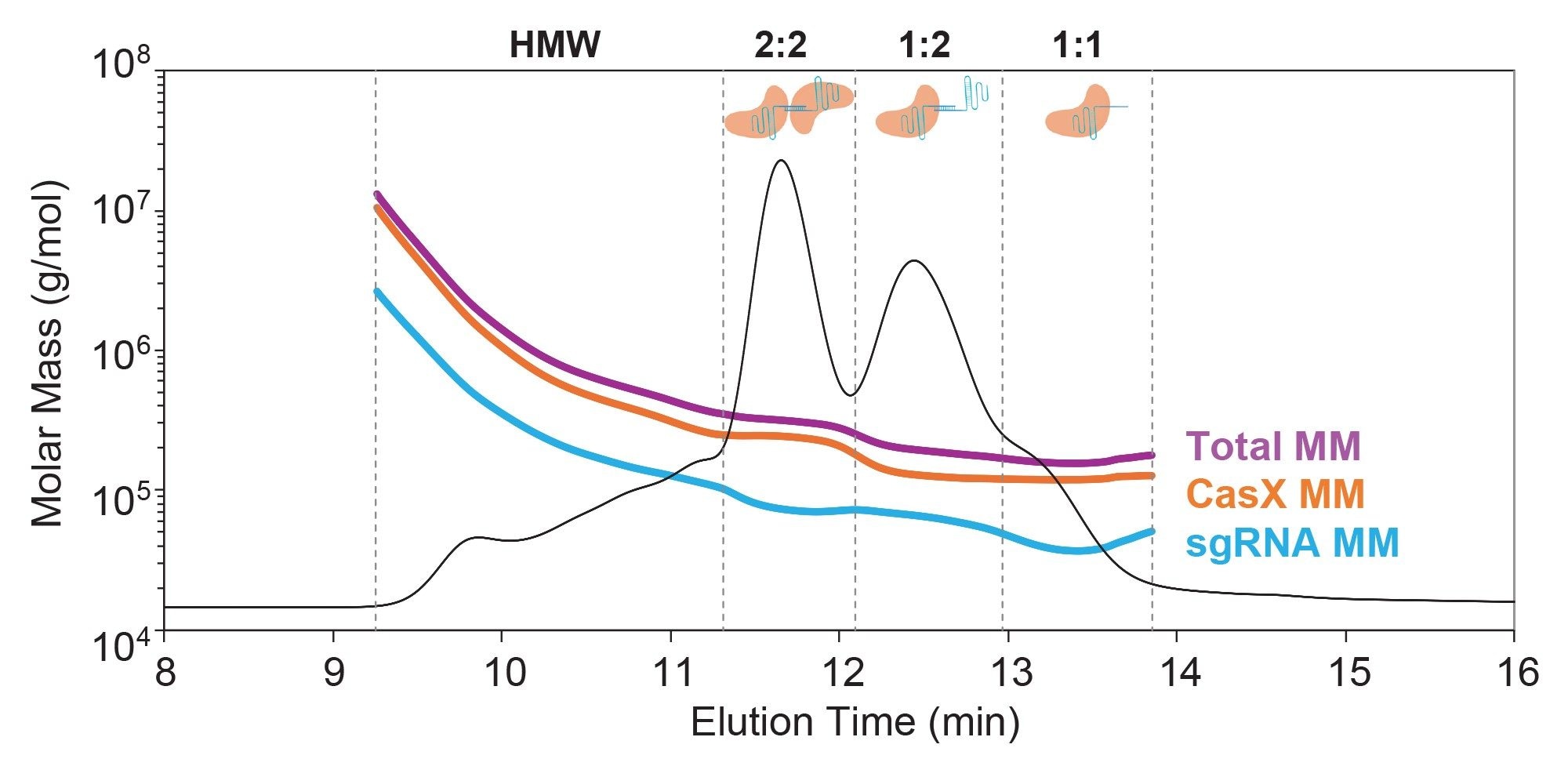
Combining SEC-MALS with simultaneous collection of UV absorbance at 260 nm and 280 nm enables compositional analysis of the RNP complex, providing direct measurement of the CasX and sgRNA molar mass at each point across the chromatogram. Analysis of the RNP complex revealed multiple coeluting peaks, each containing varying stoichiometric ratios of CasX and sgRNA (Figure 4). These data are consistent with multiple association states.
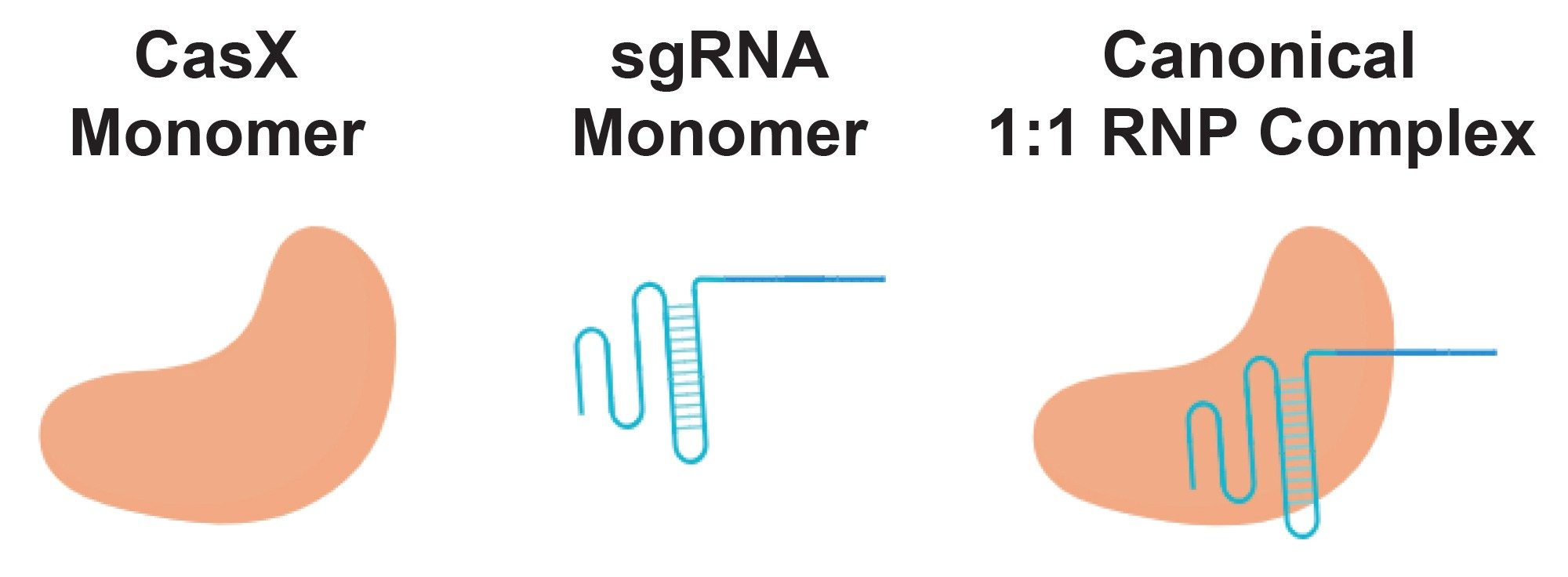
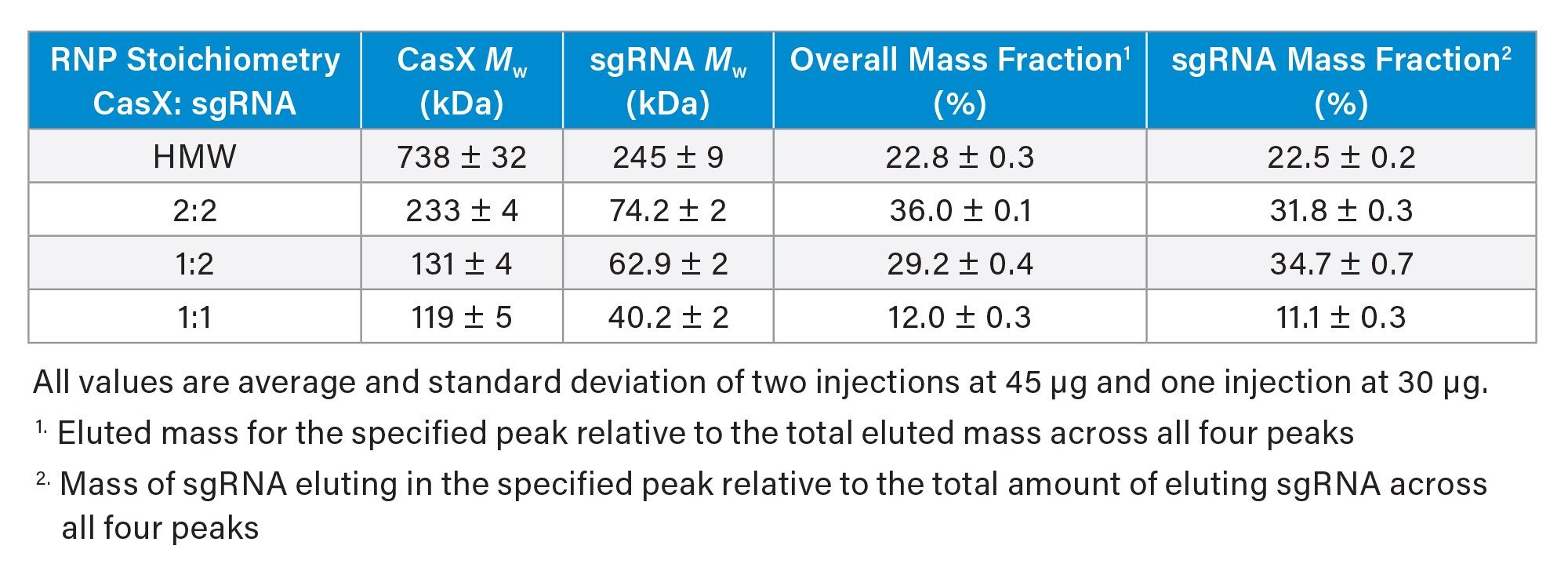
Prior structural and biochemical data suggest that the canonical RNP complex comprises one CasX monomer bound to one sgRNA monomer (Figure 1). With SEC-MALS, the presence of this 1:1 binary complex was confirmed; however, it was the least abundant in the elution profile (Table 2). Instead, the most abundant species was consistent with a dimer of sgRNA bound to two CasX proteins (~36% by mass) followed by a single CasX protein bound to two sgRNA (29% by mass). Notably, the distribution of sgRNA monomer and dimer in the RNP complex closely reflects the distribution observed in the apo sgRNA sample (66% sgRNA dimer in RNP complex vs 68% sgRNA dimer observed in apo condition).
These larger, unexpected stoichiometries likely arise from sgRNA dimers and other HOS identified in the starting material and is in line with previous studies that demonstrate that sgRNA HOS contributes to RNP heterogeneity.7 Although the CasX protein only has one binding site for sgRNA, it maintains a high affinity for nucleic acid in order to bind and cleave target DNA.8 The current hypothesis is that the sgRNA dimers and higher order species may present two or more binding sites to which the CasX monomer can bind. Since the CasX was found to be monomeric by SEC-MALS with no evidence of reversible self-association, it is unlikely that the formation of HOS is mediated by the oligomerization of CasX.
In addition to these three stoichiometries, a heterogeneous population of HMW species was also evident. Analysis of the weight fraction indicated it was predominantly protein (~80% protein and ~20% nucleic acid) and this distribution was heterogeneous across the peak indicating mixtures of coeluting species. Although the exact structure of the species is unknown in this region, it is predicted to be CasX bound to various aggregated forms of sgRNA. Detection of these undesirable species is critical for improving RNP activity.
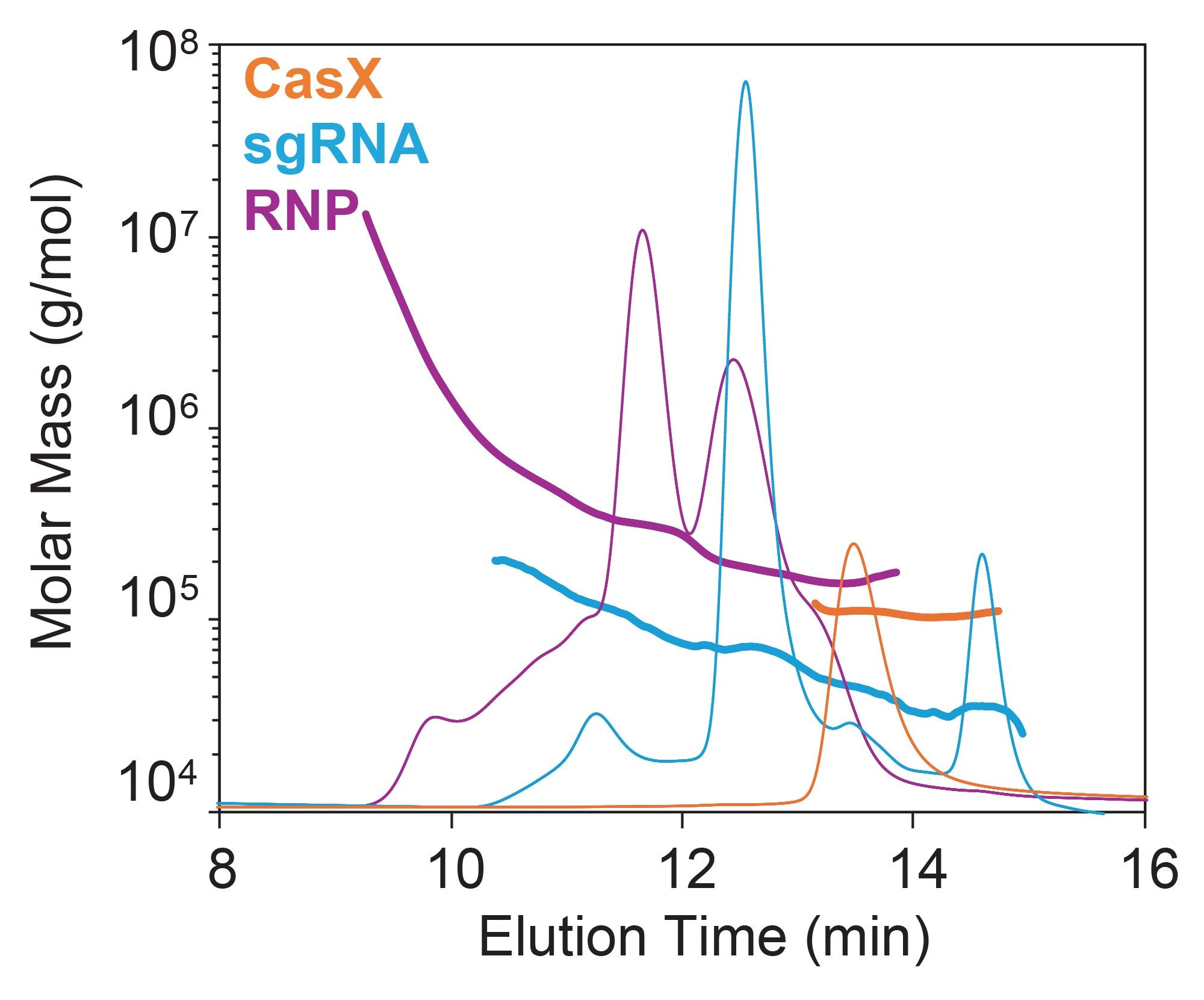
For both the RNP and the individual components, it is important to note that that retention time alone cannot be reliably used for peak identification (Figure 5). For example, the sgRNA dimer with molar mass ~70 kDa (blue chromatogram) elutes at the same time (~12.5 minutes) as the 1:2 CasX-sgRNA complex with nearly twice the molar mass (~130 kDa, purple chromatogram), suggesting significant conformational differences between pure protein, RNA oligomers, and RNP complexes. This data underscores the importance of adding MALS to SEC for positive identity of each eluting species. Moreover, the ADC Analysis Module enables direct measurement of composition, providing accuracy in the measured concentration which can be applied throughout product development.
Conclusion
In this study, key attributes, including oligomeric state and purity, were evaluated for CRISPR/Cas components using SEC-MALS. The SEC-MALS results identified dimers and other HOS present in the sgRNA starting material. When the SEC-MALS method was combined with multiple concentration detectors (UV260 and UV280 nm and dRI), this approach enabled identification of absolute stoichiometry of the RNP sample. Additionally, the SEC-MALS data revealed the presence of multiple RNP complexes with the most abundant complexes containing a non-stoichiometric number of sgRNA molecules. These eluting complexes positively correlated with the distribution of sgRNA in the apo sample, providing deeper insight into the heterogeneity of these molecules. The SEC-MALS method presented here may support researchers in identifying sgRNA HOS and improving the formation and activity of RNP complexes.
References
- Jinek, M.; et al. A Programmable Dual RNA-Guided DNA Endonuclease in Adaptive Bacterial Immunity. Science 2012, 337 (6096), 816–821. https://doi.org/10.1126/science.1225829.
- Jiang, F.; Doudna, J. A. CRISPR–Cas9 Structures and Mechanisms. Annu. Rev. Biophys. 2017, 46 (Volume 46, 2017), 505–529. https://doi.org/10.1146/annurev-biophys-062215–010822.
- Commissioner, O. of the. FDA Approves First Gene Therapies to Treat Patients with Sickle Cell Disease. FDA. https://www.fda.gov/news-events/press-announcements/fda-approves-first-gene-therapies-treat-patients-sickle-cell-disease (accessed 2025-07-23).
- Exagamglogene Autotemcel for Severe Sickle Cell Disease | New England Journal of Medicine. https://www.nejm.org/doi/full/10.1056/NEJMoa2309676 (accessed 2026-01-08).
- Jung, W. J.; Park, S.-J.; Cha, S.; Kim, K. Factors Affecting the Cleavage Efficiency of the CRISPR-Cas9 System. Anim. Cells Syst. 28 (1), 75–83. https://doi.org/10.1080/19768354.2024.2322054.
- Farzan, M.; Ross, A.; Müller, C.; Allmendinger, A. Liquid Crystal Phase Formation and Non-Newtonian Behavior of Oligonucleotide Formulations. Eur. J. Pharm. Biopharm. 2022, 181, 270–281. https://doi.org/10.1016/j.ejpb.2022.11.021.
- Camperi, J.; Moshref, M.; Dai, L.; Lee, H. Y. Physicochemical and Functional Characterization of Differential CRISPR-Cas9 Ribonucleoprotein Complexes. Anal. Chem. 2022, 94 (2), 1432–1440. https://doi.org/10.1021/acs.analchem.1c04795.
- Liu, J.-J.; et al. CasX Enzymes Comprise a Distinct Family of RNA-Guided Genome Editors. Nature 2019, 566 (7743), 218–223.
Featured Products
720009402, June 2026