SARS-CoV-2 コロナウイルスが原因で発生した最近の新型コロナウイルス感染症パンデミックのアウトブレイクにより、現在までに数百万人が罹患し、世界の死者数は 200 万人を超えています1。このウイルスは比較的新しいことから、科学コミュニティは、有効なワクチンと免疫療法の開発を支援するため、ウイルスの生物物理学的特性の迅速な特性解析と知見の集積に取り組んできました。SARS-CoV-2 のウイルス粒子は、突起状の膜貫通型スパイクタンパク質で、宿主細胞の ACE2(アンジオテンシン変換酵素 2)受容体に結合することで、細胞に侵入します2。 SARS-CoV-2 コロナウイルスのスパイクタンパク質は、2 つの高度にグリコシル化されたサブユニット S1 と S2 で構成されるホモ三量体のクラス I 融合タンパク質です3,4。 インフルエンザなどの既知のウイルス性病原体のこれまでの研究では、外部被覆の糖鎖組成が、特に免疫認識部位の立体障害によって、免疫の回避に重要な役割を果たすことが明らかになっています5-7。
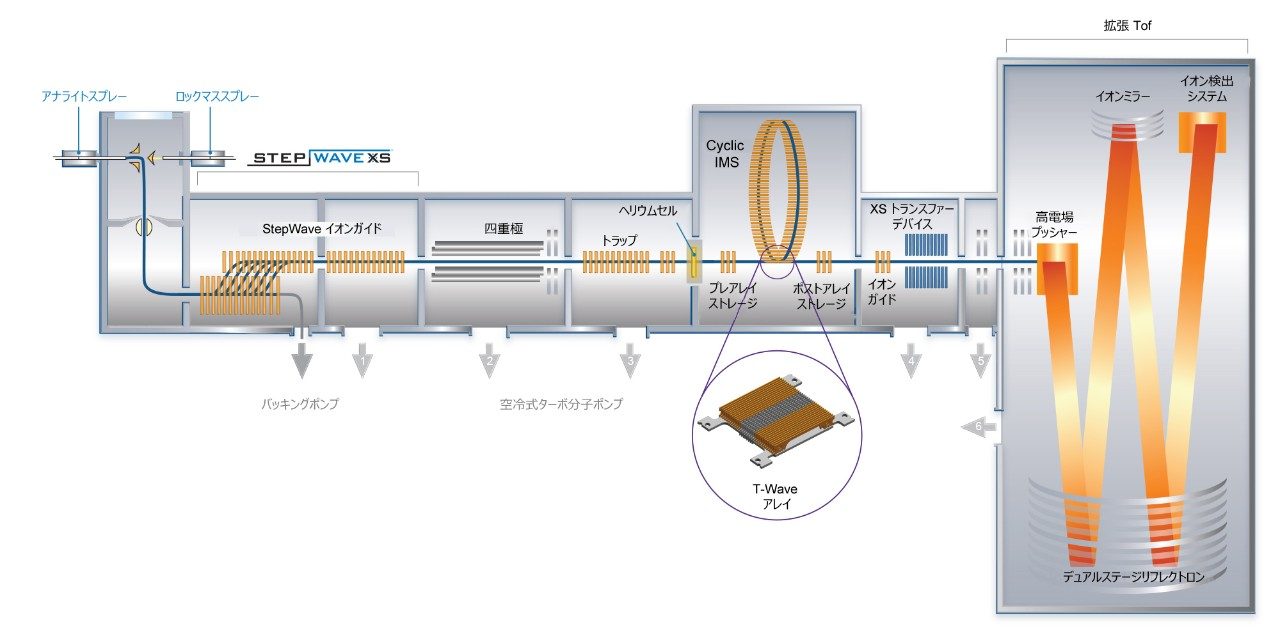
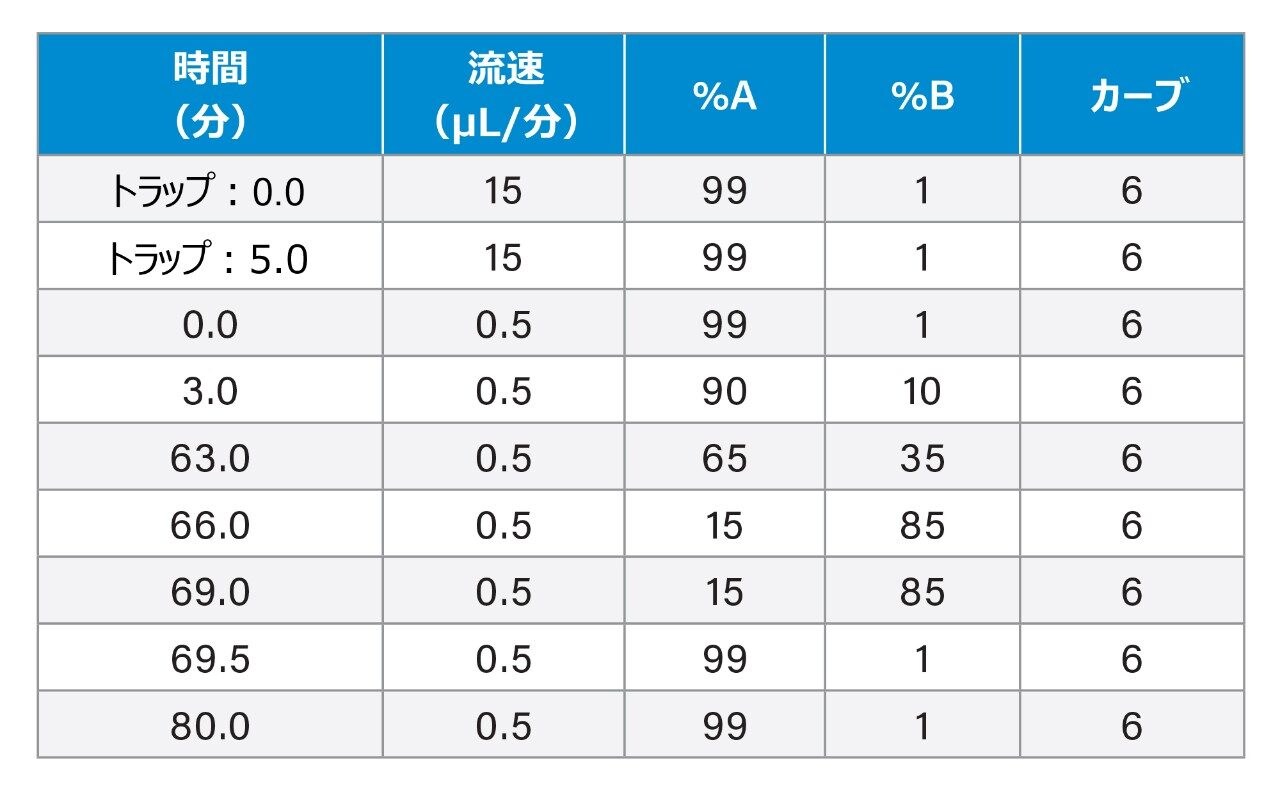
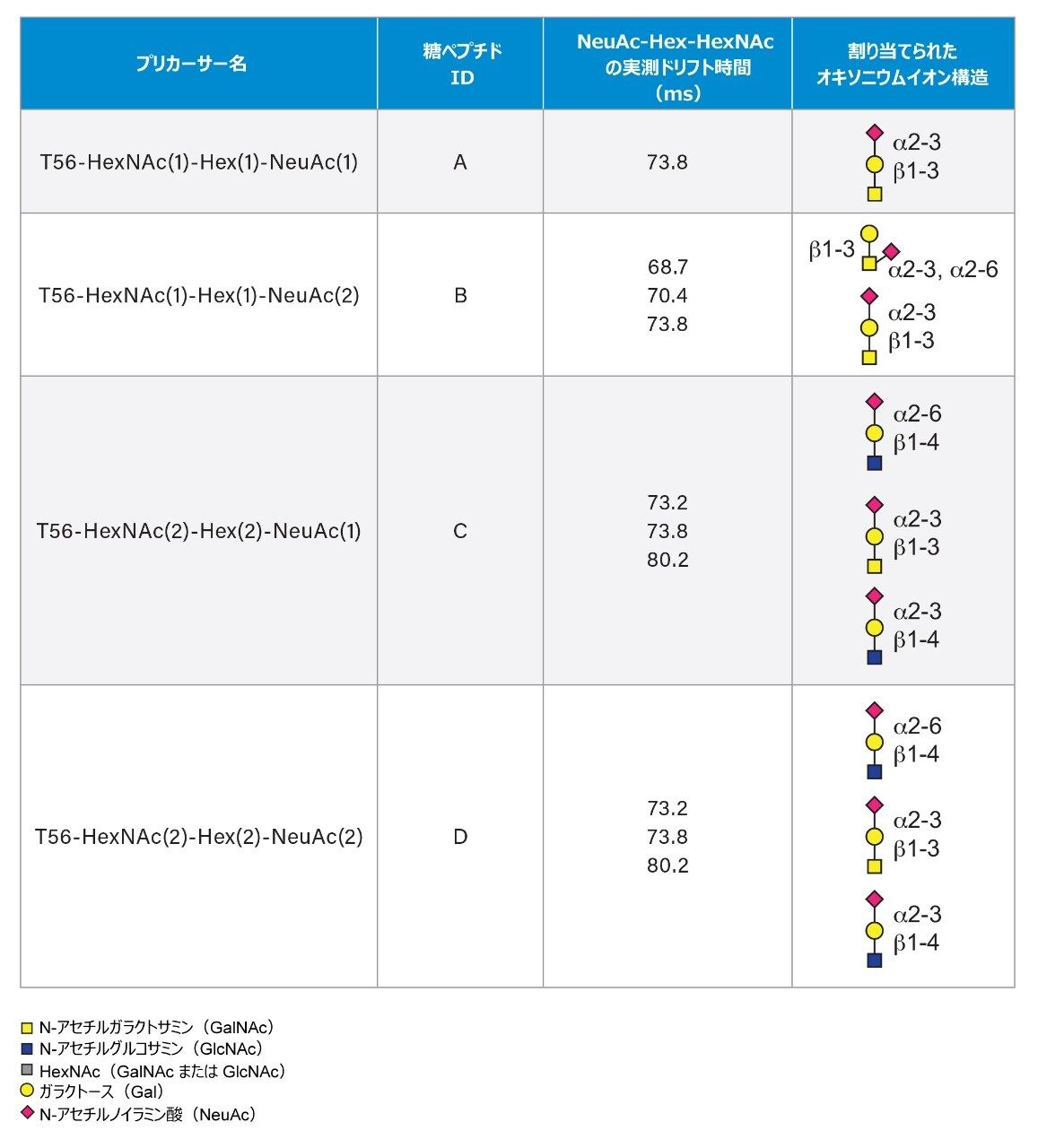
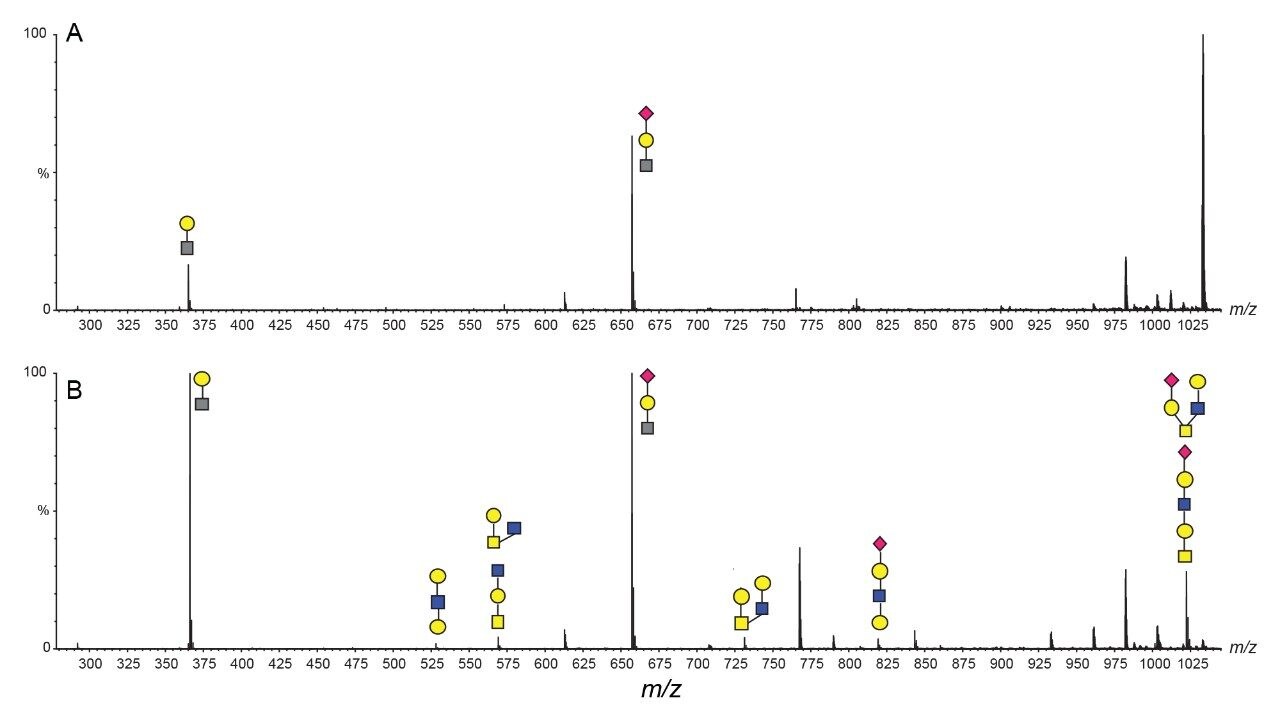
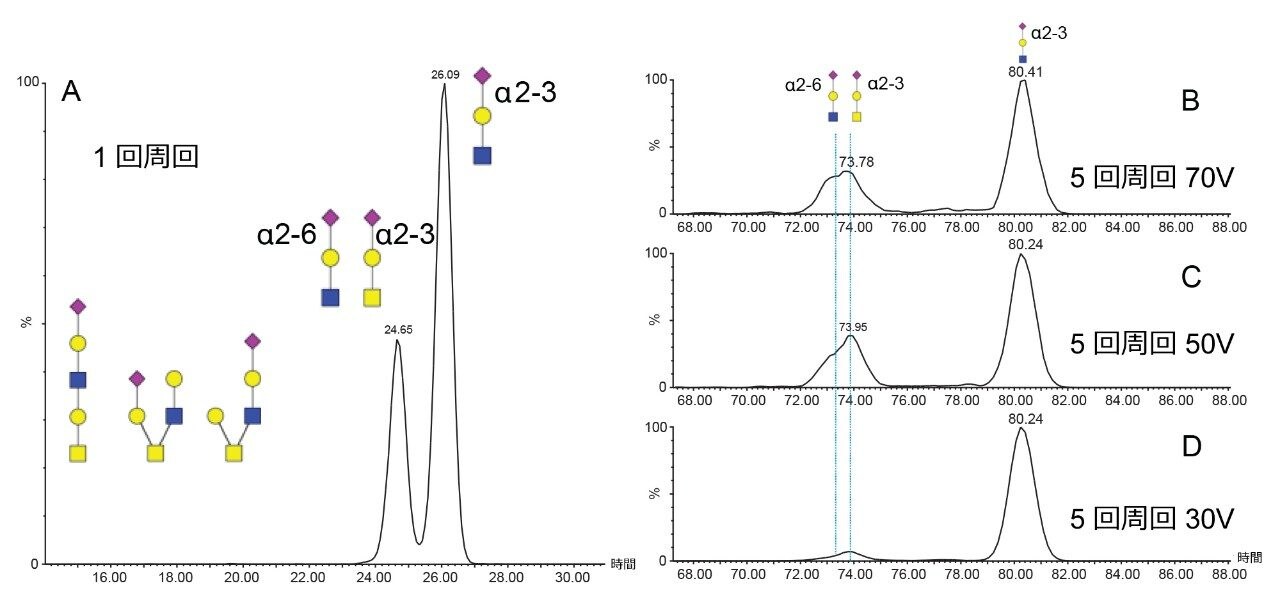
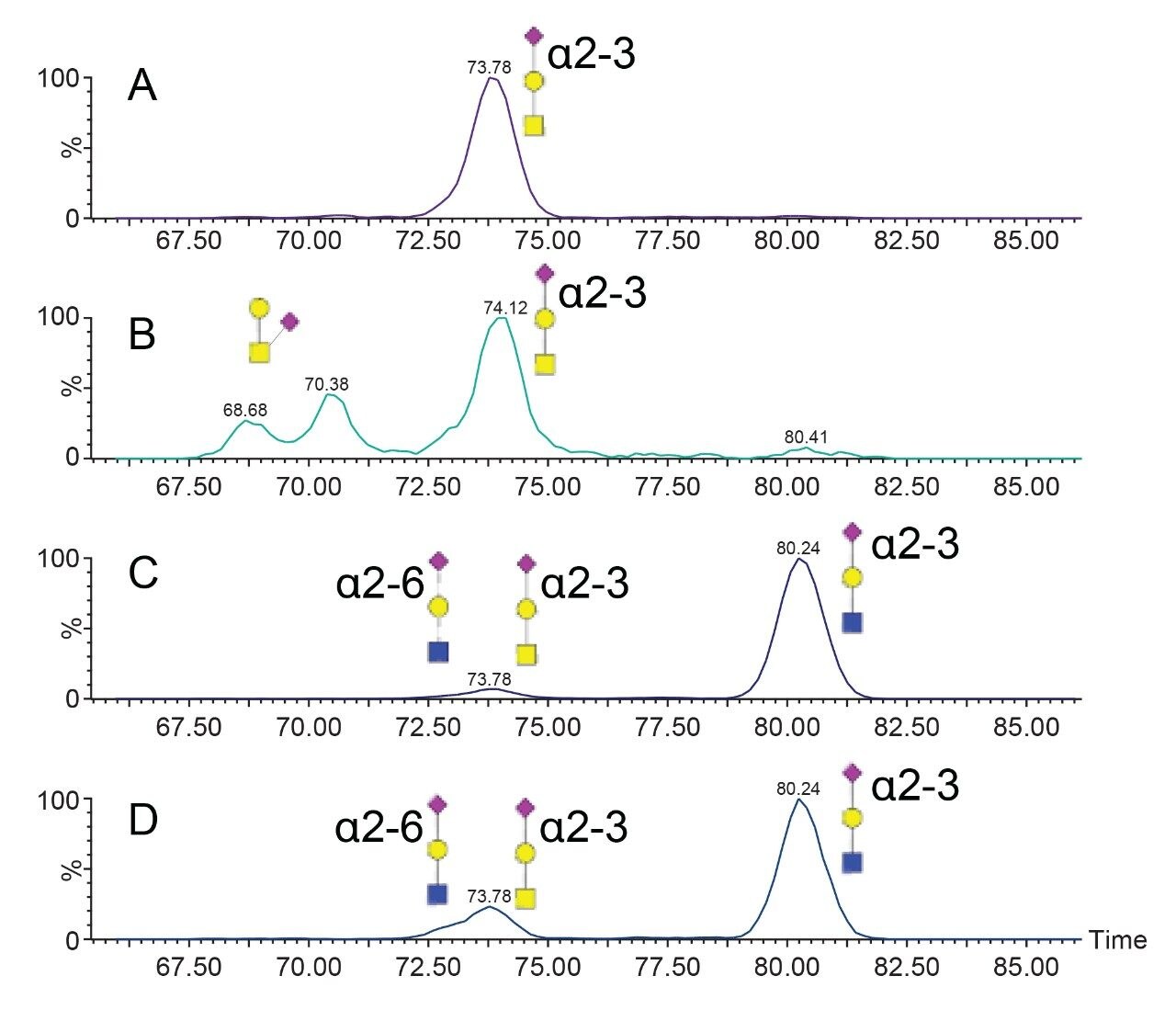
SARS-CoV-2 病原体の特性解析の取り組みには、コロナウイルスのスパイクタンパク質のグリコシル化に関する包括的な研究が含まれています。これまで、これらの研究により、利用できる 22 箇所の N 型グリコシル化部位のうち 14 の部位がコンセンサス結合しており、残りの 7 部位が相反結合していることが示されています8-16。 さらに、Shajahan らは、S1 ドメインに 3 つの O 型グリコシル化部位があることを示す証拠を示しており、そのうちの 1 つが Sanda らによって確認されました10,11。Andersen らは、SARS-COV のスパイクタンパク質のシーケンス解析の結果、S1 サブユニットと S2 サブユニットに隣接するリンカー領域に、タンパク質活性化に不可欠なフーリン切断部位が存在することを予測しました2。フーリン切断部位を含むこの領域には、さらに最大 3 箇所の O 型グリコシル化部位の存在が予測されましたが、この機能は感染性と伝染性に関連していると推測されています2。 O 型糖鎖の特性解析は、コンセンサス配列がない、グリコフォーム間の不均一性が大きい、相対存在量が低いなど、複数の理由から、非常に困難であることが知られています。そのため、これらの種類の分析種の特性解析には、高速クロマトグラフィーおよび高分解能で高感度の質量分析が必要になります。Waters SYNAPT プラットホームを用いた以前の研究では、分岐型および線状グリカン構造の異性体グリカンを分離し、α2-3 および α2-6 結合の異性体をグリカン標準品から分離する能力が示されています12–16。SELECT SERIES Cyclic IMS による MS/MS フラグメンテーションにより、塩基性アミノ酸が多いフーリン切断部位の前にあるリンカー領域に沿った O 型グリコシル化の明確なエビデンスが得られ、サイクリックイオンモビリティーを使用することで、O 型糖鎖構造を部位特異的に分離し、特性解析を行うことができます。このユニークな実験によって得られたスケーラブルな分離能は、コア 1、伸展コア 1、およびコア 2 構造の混在のエビデンスを提供し、NeuAcα2-3Galβ1-3 GalNAc および NeuAcα2-6Galβ1-4GlcNAc、非常に類似した衝突断面積を持つ異性体の分離を達成しています。